БОТАНІКА З ОСНОВАМИ ГІДРОБОТАНІКИ (ВОДНІ РОСЛИНИ УКРАЇНИ) - Б.Є. Якубенко - 2011
VI. СИСТЕМА РОСЛИННОГО СВІТУ
НИЖЧІ СПОРОВІ РОСЛИНИ
ЗАГАЛЬНА ХАРАКТЕРИСТИКА ТА ОСОБЛИВОСТІ
Відділ Бурі водорості - Phaeophyta
Еукаріотичні, багатоклітинні, переважно макроскопічні (до 60 м), морські водорос- ті, нитчастого, гетеротрихального та псевдо- чи справжньотканинного типу структури слані, з листоподібними, стеблоподібними та ризоїдоподібними утворами, зазвичай прикріплені до субстрату ризоїдами або дископодібною підошвою, і налічують нині близько 2000 видів. Появилися вони на Землі 400-570 млн років тому (силур-девон). За своєю будовою та загальнобіологічною організацією належать до «жовтої» пігментної групи та близькоспорідненІ з жовтозеленими водоростями. Вони містять хлорофіли α та с1, с2, β- та ε-каротини, І ксантофіли: 1) жовтої пігментної групи - фукоксантин, діатоксантин, діадіноксантин, та 2) лютеїнового ряду - віолоксантин, зеаксантин, антероксантин, неоксантин.
У результаті накопичення у великій кількості каротиноїдів - фукоксантину та віолоксантину, спостерігається «маскування» хлорофілу І хлоропласти набувають яскраво-жовтого або бурого забарвлення. Хлоропласти пристінні, численні, дрібні.
Кожний хлоропласт покритий чотирма мембранами, з яких дві зовнішні утворюють хлоропластну ендоплазматичну сітку, що переходить у ядерну оболонку, а дві внутрішні є власне оболонкою хлоропласта. Між зовнішніми та внутрішніми мембранами є перипластидний простір. Пластиди вторинно симбіотичні. Тилакоїди розміщуються переважно по три, на периферії знаходиться оперізуюча ламела.
У багатьох бурих водоростей у хлоропласті є піреноїд, нерідко рудиментарний, який не пронизаний тилакоїдами, а ламінарин відкладається навколо нього. Продукт асиміляції - розчинні в клітинному соці вуглеводи: ламінарин (полісахарид), ліпіди та манітол.
Манітол - шестивуглецевий вуглевод, частка якого може становити до 20-30 % сухої маси водорості та відкладається поза межами хлоропластів. Низькомолекулярні сполуки: манітол, сахароза та гліцерол слугують для запобігання замерзанню водоростей у холодних морях, а також стабілізують осмотичний тиск клітини, виконуючи осморегулюючу функцію.
Клітинна оболонка, складається з двох шарів - зовнішнього, до складу якого входять пектинові речовини, білкові сполуки альгінової кислоти і її солей (нерозчинний альгінат кальцію) та внутрішнього - целюлозного, що має фібрилярну структуру, але целюлоза відрізняється від целюлози квіткових рослин і її називають альгулозою. Більшість представників містять альгін - розчинна натрієва сіль альгінової кислоти, як і інші солі одновалентних металів. Клітини з’єднані плазмодесмами. Є пори. У цитоплазмі клітин присутні дрібні вакуолі, з яких надто дрібні - фізоди, заповнені фукозаном (флоротаніном) - речовиною, подібною до таніну. Флоротаніни відомі лише для бурих водоростей і відіграють, очевидно, захистну функцію як радіопротектори, адсорбуючу - щодо тяжких металів та охоронну - від поїдання тваринами та обростання іншими водоростями. Ядро одне, інколи у деяких представників їх багато.
Розмножуються бурі водорості нестатевим та статевим способом. Нестатеве розмноження - зооспорами та апланоспорами - тетраспорами, а також фрагментацією таломів та вивідковими бруньками. Зооспорангії бувають одногніздими чи однокамерними та багатогніздими. Статевий процес ізо-, гетеро- й оогамний. Жіночі статеві клітини продукують низькомолекулярні речовини - феромони, якими приваблюють чоловічі гамети. Гамети утворюються у багатогніздих гаметангіях, по одній у кожній камері. Зооспори і гамети грушоподібні з одним хлоропластом і двома нерівновеликими джгутиками, які розташовані на черевному боці клітини. Один джгутик довгий пірчастий, з мастигонемами, спрямований вперед, інший - коротший гладенький, спрямований назад, тобто вони гетероконтні і гетероморфні. Стигма, яка властива жгутиковим стадіям, виконує функцію лінзи, що фокусує світло на вздуття джгутика, яке і є фоторецептором.
Життєвий цикл у бурих водоростей гапло-диплофазний з чергуванням поколінь (ізоморфним або гетероморфним) або диплофазний без зміни Для представників з гапло-диплофазним життєвим циклом характерний диплоїдний спорофіт, в одногніздих спорангіях якого відбувається мейоз, і утворюються гаплоїдні зооспори та тетраспори. Вони проростають у гаплоїдні гаметофіти, які формують статеві утвори (гаметангії) з великою кількістю дрібних клітин - мікрогамети та значно меншої кількості великих макрогамет. Після копуляції зигота без періоду спокою розвивається у новий спорофіт. Якщо чергування поколінь гетероморфне, то гаметофіт, як правило, мікроскопічний, а спорофіт - макроскопічний (рис. 18).
Представники з диплофазним життєвим циклом мають диплоїдний спорофіт, проте позбавлені зооспор, мейоз відбувається під час утворення гамет, а статевий процес переважно оогамний. Оогонії та антеридії утворюються на спеціальних плідних гілочках (рецептакулах) в напівзамкнутих порожнинах - скафідіях. Зигота, яка сформувалась після запліднення яйцеклітини за межами оогонію, без періоду спокою проростає не виходячи за межі скафідію та дає початок новому поколінню.
Бурі водорості надзвичайно поширені в морях приполярних та помірних регіонів, у літоральних і субліторальних прибережних ділянках, де створюють суцільні зарості. У тропічних регіонах вони набувають інтенсивного розвитку в зимовий період (наприклад, види роду Sargassum, які утворюють значну біомасу в Саргасовому морі), за умов заниженого температурного режиму. Прісноводні форми (види родів Heribaudiella, Sphacelaria, Pseudobodanella, Lithoderma, Pleurocladia та Porterinema) є рідкісними та нараховують незначну кількість видів (близько 10). Відомі також ендофітні представники, які зростають всередині тканин інших водоростей.
Практичне використання бурих водоростей обумовлене особливостями їхнього розвитку та росту (гігантські розміри та швидке накопичення біомаси), а також специфікою хімічного складу талому. Основними напрямками практичного використання їх є отримання альгінової кислоти та альгінатів (хімічна, харчова, текстильна та медична промисловість), вирощування, збір і переробка їстівних представників, застосування біомаси як органічних добрив, багатих на калій та азот. Викиди морських водоростей у приморських країнах, як сировину багату калієм та азотом, використовують для удобрення ґрунту та на корм худобі. Основними об’єктами практичного використання є види родів Laminaria, Macrocystis, Fucus, Sargassum, Cystoseira, тощо. Проте, провідними об’єктами аквакультури є види родів Laminaria (L. saccharina, L. japonica, L. digitata) та Macrocystis (M. pyrifera), які реалізують під загальною назвою «морська капуста ».
Сучасна система відділу Phaeophyta базується на поєднанні результатів класичних методів мофолого-анатомічних досліджень (тип організації та характер росту талому, тип статевого процесу та особливості розмноження і життєвого циклу, а також наявність зооспор та піреноїду і його будова), а також сучасного молекулярно-біологічного вивчення типових представників за аналізом нуклеотидних послідовностей різних генів.
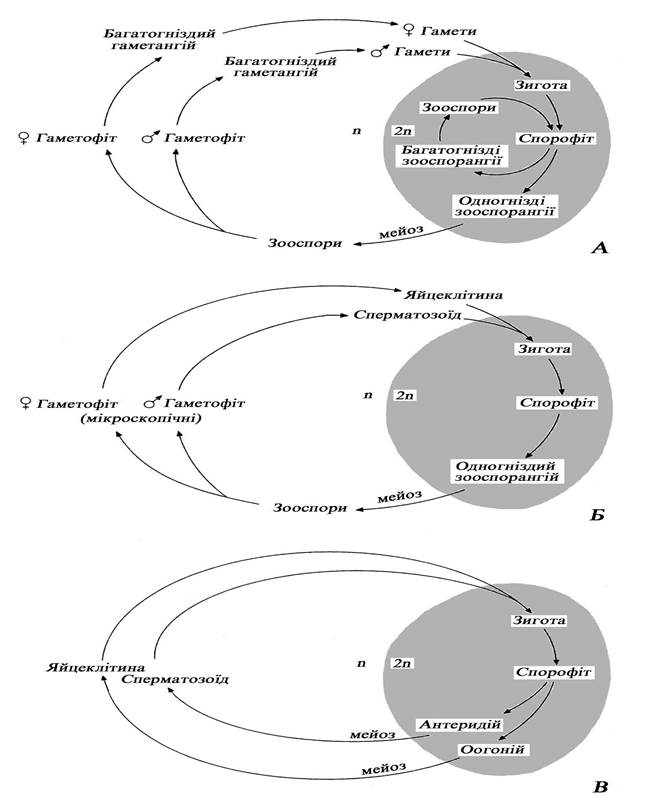
Рис. 18. Схема циклів розвитку деяких представників відділу Phaeophyta:
А - Ectocarpus (ізоморфна зміна поколінь), Б - Laminaria (анізоморфна зміна
поколінь), В - Fucus (без зміни поколінь).
За результатами цих аналізів розглядають один клас феофітових (Phaeo- phyceae), який підрозділяють на сім (інколи до 17) порядків (Ectocarpales, Scytothamniales, Cutleriales, Dictyothales, Laminariales, Sphacellariales, Fucales, тощо). Широко відомими є представники Laminariales та Fucales.