ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 1. ОСНОВИ ЗАГАЛЬНОЇ ЦИТОЛОГІЇ
1.2 Структурні компоненти клітин
1.2.4. Репродукція клітин
Одне із положень клітинної теорії стверджує, що збільшення кількості клітин, їх розмноження відбувається шляхом поділу початкової клітини. Репродукція забезпечує рівномірний розподіл редуплікованого генетичного матеріалу між двома новими клітинами.
У дорослому організмі клітини тканин і органів мають неоднакову здатність до поділу. Так, зустрічаються клітини, які зберігають здатність до поділу, постійно діляться (клітини базального шару епідерміса, крипт кишечника, кровотворні клітини червоного кісткового мозку); поряд з цим зустрічаються клітини, що втратили здатність ділитися (зернисті лейкоцити крові, остеоцити кісткової тканини, нейроцити).
Період існування клітини від поділу до поділу називається клітинним циклом. Він складається з інтерфази і мітозу. Тривалість клітинного циклу залежить від виду клітин. Так в епітеліальних клітинах порожньої кишки мишей він триває від 12,5 до 19 годин. Найбільш тривалою в клітинному циклі є інтерфаза. Вона відбувається в три періоди: пресинтетичний (G1), синтетичний (S) та постсинтетичний (G2). Різні періоди клітинного циклу відрізняються один від другого вмістом у клітині білка, ДНК та РНК.
У періоді G1, відбувається посилений ріст молодої клітини, що виникла після поділу материнської за рахунок нагромадження клітинних білків.
У наступному періоді S починається синтез ДНК. Якщо колхіцином викликати пригнічення синтезу білка або і-РНК, то перехід клітини в S-період блокується. У періоді G1 синтезуються ферменти, необхідні для утворення попередників ДНК, метаболізму РНК і білка.
S-період характеризується подвоєнням кількості ДНК в ядрі (дуплікація) і відповідно відбувається подвоєння кількості хромосом. У S-періоді також відбувається подвоєння центріолей центросоми.
Постсинтетичний період G2 (премітотичний). В цьому періоді відбувається синтез і-РНК, і триває синтез рРНК . Інтенсивно синтезуються тубуліни — білки мітотичного веретена.
Поряд з цим у тканинах тварин є клітини, які ніби виходять з циклу. Це клітини GO-періоду, у них відсутній S-період, вони не піддаються поділу. Це клітини, які тимчасово або повністю перестали розмножуватися. Клітини після диференціації не втрачають здатності до поділу і при необхідності повертаються у цикл. Прикладом їх є клітини печінки, при видаленні її частини вони починають синтезувати ДНК і вступають у мітоз, відбувається її регенерація.
Мітоз
Мітоз (від лат. mitosis - нитка) або каріокінез (від гр. сагіоn - ядро, cinesis — рух) це непрямий поділ клітин, надзвичайно важливий і необхідний етап у житті, який забезпечує розмноження клітин і розподіл між новими клітинами усіх спадкових одиниць — генетичної інформації.
Розрізняють такі основні фази мітозу: профаза, метафаза, анафаза, телофаза (рис. 15).
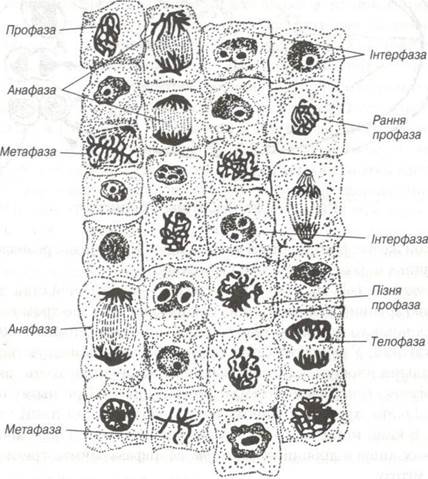
Рис. 15. Мітоз в клітинах корінця цибулі х 400
Профаза (від гр. protos - перший, phasis — явище) починається з припинення нормального функціонування клітини. При цьому вона може втрачати свої спеціальні структури (війки, тонофібрили, десмосоми) і набуває округлої форми (рис. 16).
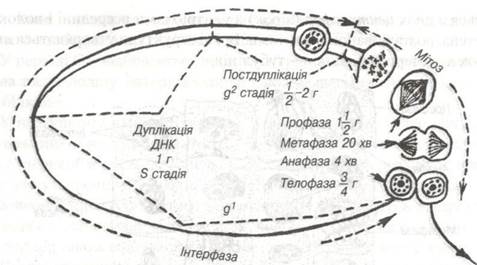
Рис. 16. Схема клітинного циклу.
Наступний етап профази - утворення мітотичного апарату. Внаслідок розходження центріолей навколо них формуються мікротрубочки. Сформований апарат поділу має веретеноподібну форму і складається з двох центросфер (зірок) з центріолями всередині і волокон веретена, розташованого між ними. Всі ці структури утворюються внаслідок полімеризації білків - тубулінів.
Інша, хроматинова частина мітотичного апарату утворюється за рахунок клітинного ядра. Комплексні сполучення молекул ДНК і білка-гістона (ДНП) інактивуються. При цьому вони скорочуються в спіраль і потовщуються, в такому стані їх можна побачити під світловим мікроскопом у вигляді грудочок хроматину. Потім вони зливаються у вигляді клубка ниток.
В наступному хроматин розпушується і в ньому можна спостерігати окремі ділянки - хромосоми.
В той же час розчиняється ядерце, ядерна мембрана розпадається. Інші органели розходяться до плазмолеми.
Метафаза (від гр. mete - після phasis — явище) цей стан клітини особливо зручний для генетичних досліджень тому, що хромосоми досягають повного розвитку. Інколи по об’єму вони займають 50% розмірів клітини. У середині метафази хромосоми вишиковуються в екваторіальній площині клітини і веретена поділу утворюють так звану метафізарну пластинку або «материнську зірку». При цьому центромерні ділянки хромосом спрямовані до центру, а їхні плечі на периферію. В кінці метафази кожна хромосома утворена з двох хроматид, з’єднаних лише в ділянці кінетохора. Метафаза займає третину часу всього мітозу
Анафаза (від гр. ana - над, зверху) характеризується двома взаємопов’язаними процесами. По-перше, мітотичне веретено ще більше витягується і клітина видовжується, по-друге обидві хроматиди кожної хромосоми внаслідок розчеплення центромери відштовхуються одна від одної і відходять кожна до свого полюсу
Кожна хроматида стає самостійною хромосомою і рухається до свого полюсу зі швидкістю 0,2-0,5 мкм/хв. Механізм руху хромосом точно не з’ясований. Більшість дослідників дотримуються гіпотези (ковзання ниток), згідно з якою сусідні мікротрубочки веретена за рахунок скоротливих білків тягнуть хромосоми до полюсів.
Телофаза (від гр. telos — кінець) після відходження хромосом до полюсів відбуваються заключні цитокінетичні та каріокінетичні процеси. Перші обумовлені виникненням течії цитоплазми з утворенням перетяжки поміж майбутніми дочірніми клітинами в ділянці колишнього екватора. Тут циркулярно розташовують дяктинові мікрофібрили. Перетяжка стає тоншою, мітотичне веретено зникає. Екваторіальна ділянка ущільнюється і утворює так зване залишкове тільце. Це тільце швидко відщеплюється і дочірні клітини остаточно відокремлюються.
Каріокінетичні процеси в телофазі начебто повторюють в зворотньому порядку всі зміни, які відбувалися в ядрі під час профази. Зокрема, хромосоми розкручуються і включаються в процеси обміну. З елементів ендоплазматичної сітки утворюється каріолема, з’являється ядерце.
Ендорепродукція - це процес утворення клітин з підвищеним вмістом ДНК (рис 19). Поява таких клітин відбувається внаслідок повної відсутності або незавершеності окремих етапів мітозу. Існує декілька моментів в процесі мітоза, блокада яких призводить до його зупинки і появи поліплоїдних клітин. Блокада може наступити при переході від С2-періода до власне мітозу або в профазі і метафазі з порушенням веретена поділу. В зв’язку з цим цитотомія не відбувається і виникають одноядерні і двохядерні поліплоїдні клітини. Поява поліплоїдних соматичних клітин може виникнути внаслідок блокади поділу. В печінці дорослих ссавців спостерігаються диплоїдні, тетраплоїдні та октаплоїдні (8n) клітини. В процесі поперемінного утворення двохядерних і одноядерних клітин з’являються ядра з 8n ,16n і навіть 32n кількістю хромосом. Таким чином з’являються поліплоїдні клітини в печінці, епітелії сечового міхура, ацинусах слинних залоз, підшлункової залозах, та пігментному шарі сітківки.
Поліплоїдні клітини функціонально активніші ніж звичайні, тому вони властиві спеціалізованим високодиференційованим тканинам.
Морфологія хромосом. Хромосоми добре забарвлюються основними барвниками. Вони помітні в ядрі клітини під час мітозу. Хромосоми не зникають після закінчення мітозу завдяки деконденсації вони набувають іншого вигляду і тому їх не видно, як окремі тільця. Як інтерфазні, так і мітотичні хромосоми складаються з елементарних хромосомних фібрил — молекул ДНП. Останнім часом вважають, що кожна хромосома побудована з однієї гігантської молекули ДНП (дезоксинуклеопротеїда), запакованої у відносно коротке тільце — власне
мітотичну хромосому. Виявлено, що у мітотичній хромосомі гігантська молекула ДНП утворює бічні петлі. Типова хромосома людини може містити 2600 петель, кожна з яких утворена ділянкою хроматинової фібрили із середньою довжиною 400 нм.
Морфологію мітотичних хромосом краще вивчати у період їх найвищої конденсації. У метафазі та на початку анафази, хромосоми у цьому стані являють собою паличкоподібні утворення різної довжини і товщини. У більшості хромосом легко знайти зону первинної перетяжки (центромери), яка ділить хромосому на два плеча. Хромосоми з рівними або майже рівними плечами називають метацентричними, а плечами різної довжини — субметацентпичними. Паличкоподібні хромосоми з дуже коротким, майже не помітним другим плечем — акроцентричні (рис. 17).
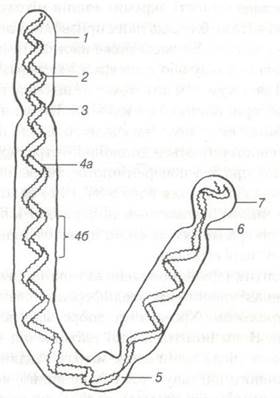
Рис. 17. Схема будови хромосоми.
1-пелікула; 2-хроматида; 3-первинна перетяжка; 4а-мала спіраль; 4б-велика спіраль; 5-центромер;
6-вторинна перетяжка; 7-супутник.
Ділянка первинної перетяжки містить кінетохор, який є центром організації мікротрубочок, що утворюють хромосомні нитки веретена поділу. У деяких хромосом є й вторинні перетяжки, які знаходяться близько одного із кінців хромосоми і відокремлюють так званий супутник хромосоми. Вторинні перетяжки ще називають ядерцевими організаторами, оскільки у цих ділянках на початку інтерфази утворюється ядерце. Кінцеві ділянки плечей називають теломерами. У тваринних організмів, як і у рослин, кількість, розміри та будова хромосом мають свою специфіку. Сукупність ознак хромосомного набору називають каріотипом. Каріотип коня, наприклад, характеризується наявністю 33 пар хромосом, серед яких 32 пари аутосоми і одна — статева хромосома. Серед останніх, які називають гоносоми, розрізняють X- та Y-хромосоми. Кількість хромосом у інших сільськогосподарських тварин також значно коливається. Кількість хромосомних наборів у клітині позначають терміном «плоїдність» і літерою n. Соматичні клітини містять диплоїдний (подвійний) набір хромосом (2 n). Статеві клітини — гаплоїдний (одинарний n), що відповідає поняттю геном, тобто сукупності ядерних елементів генетичної конституції особи. Якщо клітина має 3 n набір хромосом, її називають триплоїдною, якщо 4 n — тетраплоїдною. Велику кількість хромосомних наборів називають поліплоїдією.
Для хромосомного аналізу застосовують метод диференційного забарвлення хромосом, вперше запропонований — Касперсоном. Так, за допомогою диференційного забарвлення нефлуоресційними барвниками (суміш Гімза) вдалося виявити, що кожна хромосома забарвлюється неоднорідно вздовж її довжини, при цьому спостерігається чергування фарбованих і нефарбованих ділянок — так звана диференційна неоднорідність хромосом. Це дає змогу чітко ідентифікувати кожну хромосому, а також складати так звані хромосомні карти з визначенням локалізації у них певних генів (рис. 18).
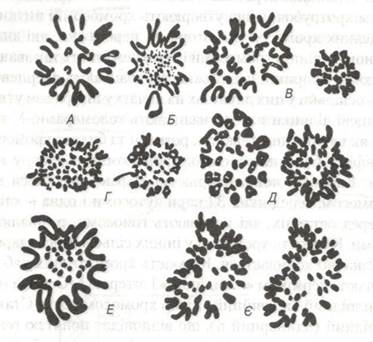
Рис. 18. Хромосоми різних видів тварин:
А - щуки (2п=18); Б - курей (2п=78); В - котів (2п=38)4 Г - коней (2п= 66); Д - бугая (2н=60); Е - саламандри (2н=34); Є - вівці (2п=54).
Амітоз - прямий поділ соматичних клітин, що відбувається без видимих під мікроскопом морфологічних перебудов в ядрі та цитоплазмі. Під час цього поділу ядро перебуває в інтерфазному стані, також не утворюється мітотичний апарат.
Амітоз починається з поділу ядерця, потім ядра і цитоплазми. Проте поділ цитоплазми відбувається не завжди, внаслідок чого утворюються дво-і багатоядерні клітини.
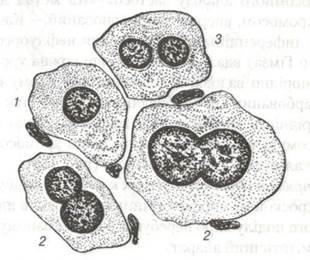
Рис. 19. Ендомітоз в клітинах десцеметової оболонки ока коня:
1-одноядерна клітина; 2-поділ ядра; 3-двоядерна клітина.
Розрізняють - генеративний, дегенеративний та реактивний амітоз.
- Ґенеративний амітоз найбільш чітко проявляється у простіших організмів. В порівнянні з мітозом він значно простіший і не супроводжується спіралізацією хромосом. Такий процес часто називають фрагментацією. Він забезпечує швидке розмноження клітин.
.Дегенеративний амітоз спостерігається у клітин, які втратили здатність до мітотичного поділу. Не маючи механізму для точного розділення молекул ДНК такий амітоз не має генеративного значення і звичайно передує відмиранню клітин.
Реактивний амітоз виникає при різкій активізації клітин в зв’язку з ушкодженням тканини. Він проявляється в масовому, швидкому поділі ядер, які також швидко руйнуються.
Отже, даний процес в багатьох випадках носить абортивний характер. Відновлення ушкодженої тканини забезпечується мітозом, який приходить на зміну реактивному прямому поділу ядер.
Приклад такого поділу буде розглянутий при описанні змін в ушкодженому нерві.