ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 1. ОСНОВИ ЗАГАЛЬНОЇ ЦИТОЛОГІЇ
1.2 Структурні компоненти клітин
1.2.3 Ядро
Ядро ( від лат. nucleus і гр. саrіоn) є однією з важливих складових частин клітини. Воно забезпечує збереження і підтримку спадкової інформації у вигляді незмінної структури ДНК. В ядрі відбувається відтворення або редуплікація молекул ДНК, що дає змогу при мітозі двом дочірнім клітинам одержувати цілком однакові в якісних і кількісних відношеннях об’єми генетичної інформації.
Другою функцією ядра є утворення власне апарата білкового синтезу. Це не лише синтез, транскрипція на молекулах ДНК різних інформаційних РНК, а й транскрипція усіх видів транспортних і рибосомних РНК. У ядрі також утворюються субодиниці рибосом шляхом комплексування синтезованих в ядерці рибосомних РНК з рибосомними білками, які синтезуються в цитоплазмі і переносяться в ядро (рис. 13).
Ядро може перебувати у мітотичному стані — під час поділу клітини та в інтерфазному — між поділами — метаболічні ядра. У живій клітині інтерфазне ядро оптично пусте, в ньому видно лише ядерце. При дії різних пошкоджуючих агентів клітина набуває стану паранекрозу. З цього стану клітина може повернутися до нормальної життєдіяльності або загинути. В ядрі у цей час морфологічно розрізняють зміни, характерні для загибелі клітини^саріопікноз — ущільнення, каріорексис — розпад, каріолізис — розчинення.
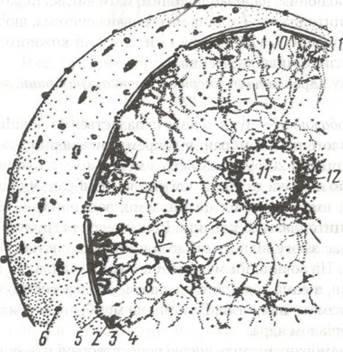
Рис. 13. Схема інтерфазного ядра за Заварзіним і Хазаровою:
1-поверхневий апарат ядра; 2-3— мембрани ядерної оболонки (2-зовнішня, 3-внутрішня; між ними перинуклеарний простір); 4-щільна пластинка; 5-комлекс з порами; 6-рибосоми; 7-гетерохроматин; 8-еухроматин (ДНП); 9-елементи ядерного матрикса; 10-РНП-частинки; 11-ядерце; 12-навколо- ядерцевий хроматин.
Таким чином, ядро являє собою одну з важливих складових частин клітини. Усі клітини тварин містять ядро, за винятком зрілих клітин крові — еритроцитів. Більшість клітин має одне ядро, винятком є двоядерні клітини (25-30%) печінки великої рогатої худоби, кролів, білих щурів і свиней. У покривному шарі перехідного епітелію зустрічаються триядерні клітини, а багатоядерні — в остеокластах кісткової тканини, а також в нервових клітинах гангліїв простати. Форма ядра різна, однак в більшості випадків вона відповідає формі клітини. Так, ядро лімфоцита кулясте, клітин гладеньких м'язів — паличкоподібне, гепатоцитів — кулясте, ядро жирових клітин під впливом жирових накопичень сплющене та відтиснене до плазмолеми. У слинних залозах, клітини яких продукують слизовий секрет, ядро також сплющене і розміщується у базальній частині клітини. Ядро лейкоцитів, крім кулястого, може бути сегментованим, бобоподібним, паличкоподібним, мати вигляд підкови.
Ядро та цитоплазма — єдина інтегрована система, що знаходиться у постійній рівновазі. Об'єм ядра і цитоплазми кожного типу клітин має своє постійне співвідношення.
До складу ядра входять: ядерна оболонка, хроматин, ядерце та каріоплазма.
Ядерна оболонка — нуклеолема складається із зовнішньої і внутрішньої біологічних мембран, відокремлених перинуклеарним простором завширшки 20-60 нм. Кожна з мембран має товщину до 8 нм і морфологічно подібна до інших клітинних мембран. Ядерна мембрана відокремлює вміст ядра від цитоплазми, зовнішня — безпосередньо контактує з цитоплазмою клітини. Вона має ряд структурних особливостей, що дає змогу віднести її до мембранної системи ендоплазматичної сітки. На зовнішній мембрані з боку гіалоплазми знаходяться полірибосоми, а сама зовнішня ядерна мембрана переходить в мембрани ендоплазматичної сітки. Внутрішня мембрана пов’язана з хромосомним матеріалом ядра.
Ядерна мембрана містить ядерні пори, утворені в результаті злиття зовнішньої та внутрішньої ядерних мембран. При цьому утворюються округлі перфорації до 90 нм. Вони заповнені складноорганізованими глобулярними та фібрилярними структурами, які разом з мембранною перфорацією утворюють комплекс пори. Він побудований з трьох рядів гранул по вісім штук у кожному ряді, діаметр гранул 25 нм. Від гранул відходять фібрилярні відростки. Фібрили, що відходять від периферійних гранул, можуть сходитися в центрі і утворювати своєрідні перегородки, так звані діафрагми пор. Розмір ядерних пор у кожного виду клітин є величиною сталою. Кількість ядерних пор залежить від метаболічної активності клітин, густина їх на поверхні нуклеолем більше у клітин з високою метаболічною активністю (рис. 14).
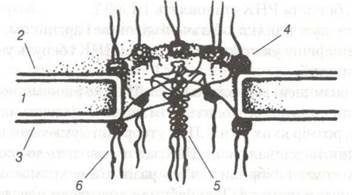
Рис. 14. Тонка організація ядерної пори:
1-перинуклеарний простір; 2-внутрішня ядерна мембрана; 3-зовнішня ядерна мембрана; 4-периферійні субодиниці;
5-центральна гранула; 6-фібрили, що відходять від гранул.
Ядерна оболонка виконує бар'єрну функцію, відокремлює вміст ядра, його генетичний матеріал від цитоплазми, обмежує вільний доступ в ядро та вихід з нього різних речовин, регулює транспорт макромолекул між ядром і цитоплазмою. Ядерна оболонка бере участь у створенні внутрішньоядерного порядку шляхом фіксації хромосомного матеріалу в інтерфазі до внутрішньої ядерної мембрани.
Хроматин (від лат. chromatunum — забарвлення). Завдяки здатності сприймати фарбники, ця складова частина інтерфазного ядра названа «хроматин» (Флемінг, 1880). Здатність хроматину сприймати лужні фарбники свідчить про його кислотні властивості, які визначаються тим, що до складу хроматину входить ДНК у комплексі з білками. Властивостями фарбуватися володіють і хромосоми, що спостерігається під час мітотичного ділення клітин. Таким чином, хроматин інтерфазних ядер являє собою хромосоми, які у цей час втрачають свою компактну форму, розпушуються, деконденсуються. Зону повної деконденсації і її ділянки мають назву еухроматин. При неповному розпушуванні хромосом в інтерфазному ядрі видно ділянки конденсованого хроматину, який називають гетерохроматином. Під час мітозу весь еухроматин конденсується і входить до його складу хромосом.
До складу хроматину входятьгскладні комплекси дезоксирибону- клеопротеїдів, що складаються з ДНК і спеціальних хромосомних білків — гістонів. Хроматин також містить РНК. В кількісному співвідношенні ДНК, білок та РНК становлять 1:1,3:0,2.
Гістони — лужні білки, збагачені лізином і аргініном. Вони забезпечують специфічну укладку хромосомної ДНК і беруть участь у регуляції транскрипції.
Гістони розміщені вздовж молекул ДНК не рівномірно, а у вигляді блоків. У один такий блок входить вісім молекул гістонів, які утворюють нуклеосоми, розмір яких 10 нм. При утворенні нуклеосом відбувається компактизація, надспіралізація ДНК, що призводить до скорочення довжини хромосомної фібрили у п'ять разів. Саме хромосомна фібрила має вигляд нитки намиста. Такі фібрили додатково повздовжньо конденсуються і утворюють основну елементарну фібрилу хроматину товщиною 25 нм. Негістонові білки інтерфазних ядер утворюють всередині ядра сітку, яку називають ядерною білковою матрицею. Вона являє собою основу, що визначає морфологію і метаболізм ядра.
Хроматин є структурним аналогом хромосом, які можна спостерігати під час поділу клітин. В деяких випадках ціла хромосома в період інтерфази може залишатися у конденсованому стані, вона має вигляд грудочки гетерохроматину. Наприклад, одна з Х-хромосом у соматичних клітинах самки залишається у конденсованому стані на стадії дроблення зиготи. Вперше цей хроматин був досліджений у ядрі клітини кішки і описаний М.Барром і Л.Бертрамом у 1949 р. Його назвали статевим хроматином, або тільцем Барра. Визначення статевого хроматину використовують для встановлення генетичної статі організму.
В ядрах, крім хроматинових ділянок і матриксу, мають місце перихроматинові фібрили, перихроматинові та інтерхроматинові гранули, які містять РНК. Матрицями для синтезу РНК є різні гени, розміщені по деконденсованих ділянках хроматинових фібрил. У хроматині міститься 1% ліпідів, роль їх не визначено.
Ядерце є найщільнішою структурою ядра (в 1.5 рази перевищує щільність ядра) і добре забарвлюється основними барвниками.
Це пов’язано з наявністю в ньому великої кількості РНК, концентрація якої тут в 2-8 разів, вища ніж у ядрі.
Кількість ядерець, як правило, відповідає кількості хромосомних наборів. Тому в диплоїдних клітинах в одному ядрі буває два ядерця.
Ядерце - це не самостійна структура, а похідне хромосом, які містять так звані ядерцеві організатори. Останні являють собою локуси хромосом з найбільш високою концентрацією і активністю синтезу РНК в інтерфазі.
Ядерце це місце утворення рибосомних РНК і самих рибосом.
Субмікроскопічна будова ядерця представлена в двох основних структурах: гранулах діаметром 15-20 нм і фібрилах товщиною 6-8 нм. Гранули є субодиницями дозріваючих рибосом. Фібрили - це рибонуклеїнові тяжі. Навколо ядерця знаходиться компактна зона навколо- ядерцевого гетерохроматина.
Каріоплазма — це рідка частина ядра, в якій містяться ядерні структури. Досліди показали, що після видалення з ядер хроматину і мембран, вони зберігають свою цілісність. Під електронним мікроскопом встановлено, що до складу каріоплазми входять комплекси пор з фібрилярним периферійним шаром, ядерцеві та численні фібрили, що знаходяться у міжхроматинових районах. Весь комплекс цих структур називають білковим ядерним матриксом. Компоненти матриксу являють собою динамічну структуру; ядерний матрикс відіграє важливу роль в підтримці загальної структури ядра, може брати участь в регуляції синтезу нуклеїнових кислот.