ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 1. ОСНОВИ ЗАГАЛЬНОЇ ЦИТОЛОГІЇ
1.2 Структурні компоненти клітин
1.2.2 Цитоплазма
До складу цитоплазми входять гіалоплазма, органели та включення.
Гіалоплазма (від гр. hyalinos — прозорий) — це прозора рідка складова частина цитоплазми або матрикс, що утворює її внутрішнє середовище. При електронній мікроскопії вона має вигляд гомогенної та тонкозернистої речовини з низькою електронною щільністю. Гіалоплазма являє собою складну колоїдну систему і містить різні біополімери: білки, нуклеїнові кислоти, полісахариди тощо. Ця система здатна переходити із рідкого стану в гель і навпаки. Окремі зони гіалоплазми можуть змінювати свій агрегатний стан залежно від умов або функціонального завдання. Деякі молекули білків — тубулінів можуть бути диспергійовані в гіалогілазмі, але в певні моменти вони починають збиратися і утворювати мікротрубочки. Таким же чином, у гіалоплазмі можуть виникати і розпадатися різноманітні фібрилярні, нитчасті комплекси білкових молекул. До ферментів матрикса відносять ферменти гліколізу, метаболізму цукрів, азотистих сполук, амінокислот, ліпідів та інших важливих сполук. У гіалоплазмі відбувається постійний рух іонів до плазматичної мембрани і від неї, до мітохондрій, ядра, вакуолі. Гіалоплазма є зоною переміщень молекул АТФ. В гіалоплазмі містяться органели та включення.
Органели — постійні мікроскопічні та ультраструктурні утворення, що виконують життєвоважливі функції клітин.
Розрізняють органели загального призначення та спеціальні, а но будові мембранні та немембранні.
Мембранні органели — це складові елементи цитоплазми, відмежовані від гіалоплазми клітинними мембранами. До них відносяться: мітохондрії, ендоплазматична сітка, лізосоми, пероксісоми, комплекс Гольджі;
Немембранні органели — рибосоми, мікрофіламенти, мікротрубочки, центросома. Всі названі вище належать до органел загального призначення. Поряд з цим в деяких клітинах існують спеціальні органели — тонофібрили епітеліальних клітин, міофібрили міоцитів та м'язових волокон, нейрофібрили нервових клітин, війки та джгутики.
Мембранні органели
Мітохондрії. Термін «мітохондрія« (від гр. mitos - нитка і chon- drion — зернятко) введено Бендою у 1897р. Під світловим мікроскопом мітохондрії мають вигляд круглих зерен та коротких паличок товщиною 0,2-2 мкм і довжиною 1-10 мкм. При розгляді під електронним мікроскопом кожна мітохондрія має овальну або видовжену форму. У них розрізняють зовнішню гладеньку та внутрішню мембрани, від останньої всередину мітохондрії відходять вирости — кристи. Проміжки між кристами заповнені матриксом — електронно-щільною речовиною, у якій виявляють ниткоподібні утворення товщиною 2-3 нм та гранули товщиною 15-20 нм. Ниткоподібні утворення матрикса являють собою молекули ДНК і РНК, а гранули — мітохондріальні рибосоми. У матриксі і у внутрішній мембрані містяться білки — ферменти, що забезпечують синтез АТФ шляхом окисного фосфорилювання аденозиндифосфату. Кількість мітохондрій в клітинах значно варіює від поодиноких до сотень. Наприклад в клітинах печінки вони складають 20% загального об’єму цитоплазми і містять в собі 30-35% загальної кількості білку. Мітохондрії концентруються у тих місцях цитоплазми, де є потреба в АТФ. Основною функцією їх є синтез ATФ, який відбувається в результаті процесів окислення органічних субстратів і фосфорилювання АДФ (рис. 7).
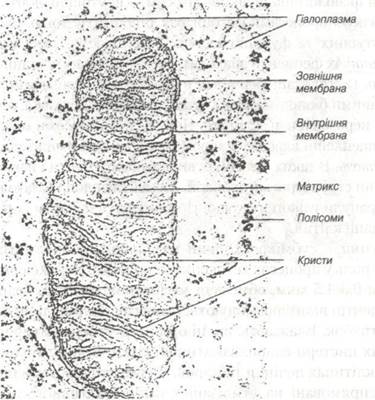
Рис. 7. Електронна мікрофотографія мітохондрії.
Мітохондрії також беруть участь у регуляції обміну води, депонування іонів Са, Mg, Р, продукції попередників стероїдних гормонів. Мітохондрії здатні рухатися; їх кількість збільшується шляхом поділу, або брунькуванням початкових мітохондрій.
Вони чутливі до різних зовнішніх впливів: голодування, дії рентгенівських променів, наркотиків тощо.
Ендоплазматична сітка (від гр. endo - в середині). Її вперше описав К. Портер у 1945 р. Вона являє собою субмікроскопічну органелу, яка утворює внутрішньоцитоплазматичну циркуляційну систему. Вона є замкненою сукупністю канальців, мішечків та цистерн, утворених безперервною біомембраною. Розрізняють два типи — гранулярну та агранулярну. Ширина канальців гранулярної сітки від 20 до 1000 нм, з боку гіалоплазми її мембрани покриті рибосомами. Останні беруть участь у синтезі білків на «експорт» і білків-ферментів, а також білків, що нагромаджуються в її цистернах та здатні транспортуватися у вакуолі комплексу Гольджі, де перетворюються і входять до складу лізосом або секреторних гранул. У ряді випадків у канальцях або вакуолях гранулярної ендоплазматичної сітки відбувається модифікація білків, зв'язування їх з цукрами і утворення секреторних гранул. Гранулярна ендоплазматична сітка здатна синтезувати мембранні інтегральні білки, які вмонтовуються в товщу мембран.
На відміну від гранулярної ендоплазматичної сітки на мембранах агранулярної ендоплазматичної сітки відсутні рибосоми. Діаметр її ка- нальців і міхурців 50-100 нм. Функція гладкої ендоплазматичної сітки пов'язана з метаболізмом ліпідів, синтезом стероїдних гормонів, глікогену, депонуванням іонів кальцію, дезактивацією отруйних речовин, що особливо характерно для гепатоцитів. Мембрана ендоплазматичної сітки безпосередньо контактує з плазмолемою клітини.
Лізосоми — мембранні органели. Виявлені і ідентифіковані в клітинах ссавців, птахів, земноводних, вони являють собою кулясті структури розміром 0,2-0,4 мкм, містять понад 60 гідролітичних ферментів, здатних до розщеплення біополімерів різної хімічної природи. Із ферментів лізосоми містять иротеїнази, нуклеази, глюкозидази, фосфатази, ліпази. Ферменти локалізуються у лізосомах і утримуються у них за допомогою ліпопротеїнової мембрани, яка обмежує і відокремлює їх вміст від зовнішніх субстратів.
Основна фізіологічна функція лізосом — внутрішньоклітинне травлення в зв'язку з процесами фагоцитозу, піноцитозу тощо. Залежно від ультраструктурних та функціональних особливостей лізосоми поділяють на первинні, їх ферменти знаходяться у неактивному стані; вторинні, або фагосоми (активовані ферменти в них безпосередньо контактують з розщеплюваними біополімерами), а також залишкові тільця, оточені біомембраною, нерозщеплені залишки. Поряд з цим лізосоми можуть брати участь у розщепленні власних макромолекулярних комплексів клітин — аутофагоцитозу. В таких лізосомах виявлені фрагменти і навіть цілі цитоплазматичні структури: мітохондрії, елементи ендоплазматичної сітки, рибосоми, гранули глікогену, тощо. Це може бути доказом їх ролі в процесах деградації клітин.
Лізосоми — субмікроскопічні мембранні органели. Відіграють вирішальну роль у процесах детоксикації клітини. Пероксісоми — тільця розміром 0,3-1,5 мкм, обмежені мембраною, містять гранулярний матрикс, в центрі якого розміщуються кристалоїдноподібні структури із фібрил і трубок. Вважають, що ці органели утворюються на розширених кінцях цистерн ендоплазматичної сітки. Вони особливо розповсюджені в клітинах печінки та нирок. Ферментні системи пероксісом (каталаза) спрямовані на утилізацію хімічно активного атомарного кисню, а також забезпечують розщеплення етилового спирту, сечової кислоти, регуляцію обміну ліпідів.
Комплекс Гольджі. Перші відомості про цю органелу належать К.Гольджі, якии в 1898р. описав її у складі нервових клітин. Під світловим мікроскопом комплекс Гольджі має вигляд сітчастого утворення. Під електронним мікроскопом він являє собою мембранні структури, що мають вигляд цистерн товщиною до 25 нм, сплющених у центральній частині і розширених на периферії. Окрему ділянку скупчення цих мембран називають діктіосомою. В проміжках між окремими цистернами знаходяться тонкі прошарки гіалоплазми. У комплексі Гольджі завершується процес формування продуктів синтетичної діяльності клітини — її кінцеве глікозування (рис. 8,9). Комплекс Гольджі бере участь у агрегації і нагромадженні продуктів, синтезованих в ендоплазматичній сітці та їх хімічній трансформації. У цистернах комплексу Гольджі синтезуються поліцукриди, які комплексуються з білками, що сприяє утворенню мукопротеїдів, а також виведенню готових секретів за межі клітини. Крім того, комплекс Гольджі забезпечує формування клітинних лізосом. (рис. 10).
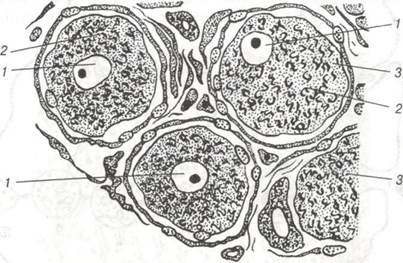
Рис. 8. Комплекс Гольджі в нервових клітинах спинального ганглію х 400: 1-ядро; 2-цитоплазма; 3-фрагмент комплексу Гольджі.
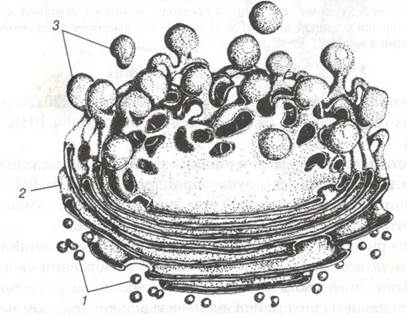
Рис. 9. Схема комплексу Гольджі:
1-транспортні міхурці; 2-зріла поверхня мішечків; 3-секреторні міхурці.
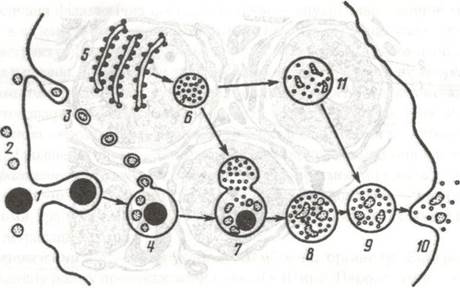
Рис. 10. Схема функціонування лізосоми і внутрішньоклітинного протеолізу за Де Дювом:
1-часточка, що піддається фагоцитозу; 2-мікромолекула; 3-мікромолекули в стані піноцитозування клітиною; 4-фагосома;
5-ергастоплазма; 6-лізосоми; 7-злиття лізосоми і фагосоми; 8-протеоліз частинок і макромолекул(9); 10-екскреція залишків протеолізу; 11-протеоліз у лізосомі з утворенням фагоцитозної вакуолі.
Немембранні органели
Рибосоми. За розміром рибосоми становлять 25x20x20 нм. До їх складу входять велика і мала субодиниці рибосомної РНК та білок (рис. 11).
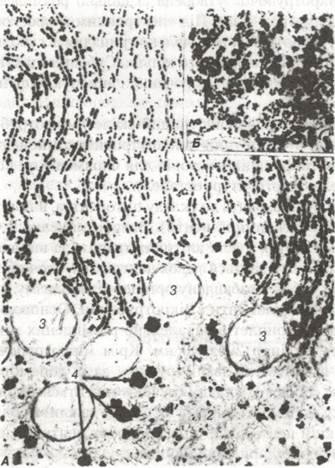
Рис. 11. Електронна мікрофотографія гранулярної ендоплазматичної сітки в цитоплазмі генатоцита щура х 38 000:
1 -паралельно розміщені цистерни, їх зовнішня поверхня містить рибосоми;
2-агранулярна ендоплазматична сітка; 3-мітохондрії; 4-глікоген.
Для стабілізації структури рибосом необхідні іони магнію. Кожна із субодиниць побудована з нуклеопротеїдного тяжа, де РНК взаємодіє з різними білками і утворює тіло рибосоми, у яких амінокислоти сполучаються, тобто синтезуються білкові молекули.
Кілька рибосом, з'єднаних спільною ниткою інформаційної РНК, називають полісомами. Останні пов’язані з мембранами ендоплазматичної сітки, синтезують білки для виведення за межі клітини. Ступінь інтенсивності синтетичної діяльності вільних рибосом менший, а утворені білки використовуються для внутрішніх потреб клітини.
Мікрофіламенти спостерігаються майже у всіх клітинах. Зокрема в рухових відростках фібробластів та мікроворсинках кишкового епітелію. Вони мають товщину 5-7 нм, побудовані з скоротливих білків: актину, міозину, тропоміозину та альфа-актину.
Розповсюджуються переважно в кортикальній зоні клітини, утворюють цитоскелет та виконують скоротливу функцію.
Вважають, що мікрофіламенти забезпечують не тільки рухомість клітини при активному амебоїдному переміщенні, але і більшість внутрішньоклітинних рухів, таких як: течія цитоплазми, рух вакуолей, мітохондрій та поділ клітин.
Мікротрубочки - являють собою прямі, не розгалуджені порожнисті циліндри діаметром 25 нм, товщиною стінки 5 нм.
Стінка мікротрубочок утворена із щільно розміщених кулястих субодиниць діаметром 5 нм. Під електронним мікроскопом на поперечних зрізах мікротрубочок виявлено тринадцять субодиниць, розташованих у вигляді одношарового кільця. Субодиниці утворені з білка тубуліну. Вони не здатні до скорочення і виконують функцію каркасних структур.
Утворюючи внутрішньоклітинний скелет, мікротрубочки можуть бути факторами руху клітини та її внутрішніх компонентів. Вони можуть виконувати функцію векторів для спрямованих потоків різних речовин та великих структур (органел).
Мікротрубочки входять до складу цитоцентра, війок та джгутиків.
Центросома (клітинний центр). Вперше описана В.Флемінгом у 1875 р. Вона міститься у всіх клітинах тварин, за винятком яйцеклітин. Центросома складається з двох центріолей, оточених центросферою, що являє собою позбавлену органел гіалоплазму, яку радіально пронизують мікрофіламенти і мікротрубочки. Основою будови центріолей є дев'ять триплетів паралельно розміщених мікротрубочок, які формують циліндр 200x500 нм. Крім мікротрубочок, до складу центріолі входять так звані «ручки», за їх допомогою триплети пов’язані між собою. У складі «ручок» міститься білок динеін, що має АТФ-азну активність і якому належить важлива роль у механізмі рухових функцій центріолей. Довгі осі обох центріолей розміщені у взаємно перпендикулярних площинах.
Перед поділом клітини центріолі подвоюються (настає дуплікація) з подальшим розходженням кожної новоутвореної пари до полюсів клітини. Дві розміщені поряд центріолі називають диплосомою. Вважається, що центріолі беруть участь в індукції полімерізації тубулінів при утворенні мікротрубочок. Центросома забезпечує розходження хромосом при поділі клітини.
Спеціальні органели
Війки та джгутики - є спеціальними органелами руху. Дійки розташовані на апікальному полюсі епітеліальних клітин дихальних шляхів і маткових труб, а джгутики входять до складу сперматозоїдів.
Довжина війок досягає 5-10 мкм, джгутиків до 150 мкм. Вони утворені виростами цитоплазми всередині яких є осьова нитка - аксонема.
Аксонема має циліндричну форму і утворена дев’ятьма дуплетами мікротрубочок і двома центральними. Базальне тільце по будові подібно до центріолі і структурно зв’язане з аксонемою в єдину систему.
Основні білки війок це тубулін та динегн. Останньому притаманна АТФ-азна активність. Війки рухаються різноманітно: маятникоподібно, хвильоподібно, лійкоподібно тощо.
Будова джгутиків описана в курсі ембріології.
Мікроворсинки це вирости цитоплазми, всередині яких знаходяться скоротливі мікрофіламенти. Останні збільшують апікальну поверхню клітини. Ця органела добре розвинена в епітеліоцитах слизової оболонки кишечника.
Тонофібрили, міофібрили і нейрофібрили. Ці різновиди мікрофібрил гіритаманні клітинам та симпластам окремих тканин. Зокрема тонофібрили та нейрофібрили відповідно епітеліальним та нервовим клітинам. Міофібрили забезпечують скорочення скелетних м’язових волокон і кардіоміоцитів.
Включення
Включення це необов’язкові компоненти цитоплазми, котрі утворюються і зникають в залежності від метаболічного стану клітини. Розрізняють: трофічні, секреторні, пігментні, вітамінні та екскреторні включення.
До трофічних включень належать жири, білки і вуглеводи. Наприклад, нейтральний жир в ліпоцитах пухкої сполучної тканини, глікоген в м’язовій тканині та клітинах печінки.
Секреторні включення - це округлі утворення різних розмірів, котрі містять біологічно-активні речовини.
Пігментні включення можуть бути екзогенного походження (каротин, барвники, часточки пилу) і ендогенного (гемоглобін, гемосидерин, білірубін, меланін, ліпофусцин, тощо). Наприклад меланоцити шкіри захищають організм від дії ультрафіолетового випромінювання. Присутність пігментів може змінювати колір клітин і тканин. Не рідко пігментація може бути діагностичною ознакою, (рис. 12).
Екскреторні включення є продуктами метаболізму, які шкідливі для клітин.
Рис. 12. Пігментні включення у меланоцитах (тотальний нефарбований препарат х 400)