ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 2. ЕМБРІОЛОГІЯ
2.2 Статеві клітини
Статеві клітини за своєю будовою подібні до соматичних клітин, але мають своєрідну будову, містять халлоїдний (половинний) набір хромосом.
Серед статевих клітин розрізняють два різновиди — сперматозоїди . та яйцеклітини — овоцити.
Сперматозоїди (від лат sperma - сімя, zoon — тварина). У різних видів сільськогосподарських та лабораторних тварин вони різняться своєю будовою. Однак для всіх них характерні морфологічні риси, що тісно пов'язані з їх основними біологічними особливостями, здатністю активно рухатися та запліднювати яйцеву клітину. При еякуляції сім'яна рідина містить мільярди сперміїв.
У сперматозоїда розрізняють голівки і хвостовії частину. Електронномікроскопічні дослідження показали, що голівка має невелике щільне ядро з гаплоїдним набором хромосом. У ядрах сперміїв містяться різні типи статевих хромосом, що несуть генетичну інформацію. Половина сперматозоїдів мають Х-хромосому, друга половина — У-хромосому. Останні хромосоми належать до аутосом. Спермії, що несуть У-хромосому при злитті з яйцеклітиною започатковують організм самця; спермії — носії Х-хромосоми при злитті з яйцеклітиною започатковують організм самки.
Передня частина головки має плоский чохлик, що вкриває акросому (від гр. akros - крайній, soma — тіло). Як чохлик, так і акросома є похідними комплексу Гольджі. В акросомі знаходяться ферменти — гіалуронідаза і протеази, які здатні розчиняти оболонку яйцеклітини та стимулювати проникнення у неї спермія при заплідненні. Зовні головка спермія покрита плазмолемою, в ній містяться фібрили, які надають їй механічної міцності. Хвостова частина сперматозоїда ниткоподібна, вона складається із шийки, проміжної, основної та кінцевої частини (рис. 20).
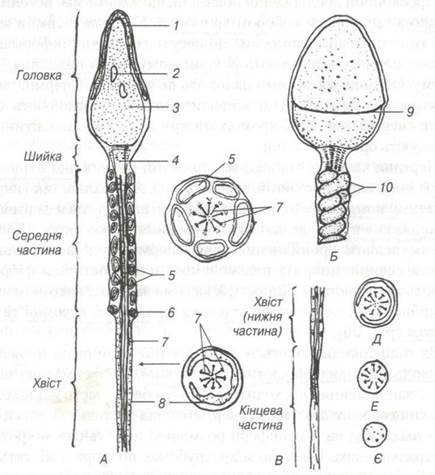
Рис. 20. Схема будови сперматозоїда самця:
1-головний чохлик; 2-вакуоль; 3-ядерний матеріал; 4-проксимальна центріоля; 5-мітохондрії; 6-каудальна частина дістальної центріолі; 7-осьові філаменти; 8-оболонка; 9-край головного чохлика; 10-мітохондріальна спіраль.
За голівкою знаходиться шийка; в ній розміщена проксимальна центріоль і краніальна частина дистальної. Проксимальна центріоль під час запліднення переходить у зиготу і бере участь у її подіті. Головна і кінцева частини хвоста сперматозоїда містить скоротливий апарат - лксонему, на її периферії розміщені дев'ять пар мікротрубочок і центральна пара. Навколо мікротрубочок по спіралі містяться мітохондрії. Мііохондрії забезпечують енергією рухливість аксонеми. У складі проміжної частини містяться циркулярні пучки мікрофібрил. Головна частина хвостика утворена аксонемою і плазмолемою з тонким прошарком цитоплазми. В останній є значна кількість циркулярно орієнтованих тонофібрилярних структур. На межі між головною і проміжною частинами хвоста навколо аксонеми знаходиться каудальна частина дистальної центріолі. Кінцева частина хвоста в своєму складі має осьову нитку з поодинокими скоротливими мікрофіламентами й покрита плазмолемою.
Біологічні властивості сперматозоїдів. В статевих шляхах самців (придаток сім'яника) у сперматозоїдів знижений обмін речовин, вони
майже нерухомі. В кінцевому відділі протоки додатка сперматозоїдів покриваються тонкою ліпоїдно-білковою оболонкою, набувають однакового електричного заряду і починають відштовхуватися один від одного.
Сперматозоїди рухаються зі швидкістю 2-5 мм за хвилину. В статевих шляхах птахів та ссавців вони рухаються прямолінійно, що залежить від симетрії голівки. Спрямування руху сперматозоїдів визначається їх реотаксисом, тобто здатністю рухатися проти течії рідини. Реотаксис проявляється лише при порівняно слабкому зустрічному потоці слизу. В кислому середовищі голівки сперматозоїдів набрякають, заокруглюються, втрачають асиметрію і тому вони набувають кругового руху, що заважає їм рухатися прямолінійно і усувається можливість зустрічі з яйцеклітинами. У сперматозоїда мало поживних речовин, тому він їх скоро втрачає і через 24-36 годин гине, якщо не зустрінеться з яйцеклітиною.
Сперматозоїди досить чутливі до іонів двох- та трьохвалентних металів, кислот, наркотиків, хініну. Глибоке охолодження до -78° або -196°С гальмує у них обмін речовин, що дає можливість продовжити термін їх зберігання на декілька років і використовувати тривалий час для запліднення.
Яйцеклітини — овоцити (від лат. ovum — яйце) — статеві клітини самки.
За своєю будовою яйцеклітини відрізняються від соматичних клітин. Зрілі яйцеклітини мають гаплоїдний набір хромосдм, після запліднення здатні розвиватися в новий організм. Вони містять більший чи менший запас поживних речовин (жовтка), необхідних для забезпечення зародкового розвитку організму, мають кортикальний шар цитоплазму. обмежені спеціальними оболонками, які захищають зародок від шкідливої дії зовнішнього середовища. Для овоцитів характерна поляризована будова за рахунок нерівномірного розміщення поживних речовин.
Поживні речовини (жовток) мають вигляд гранул або складніших утворень, що мають вигляд шарів та платівок, утворених фосфоліпідами, вуглеводами і проте)'нами Структурною одиницею жовтжа екбмілекс лігювітеліну і фосфовітину. Кожна платівка складається з щільнішої центральної і пухкої периферійної зони, зовні обмеженої осміофільною мембраною. Щільна зона утворена молекулами фосфовітина і має вигляд кристалічної сітки. Жовток синтезується при безпосередній участі ендоплазматичної сітки та комплексу Гольджі.
Особливостями овоцитів є те, що вони містять значні запаси рибосом, іРНКт тРНК. кількість яких в сотні разів перевищує їх в соматичних клітинах. В цитоплазмі яйцеклітини нагромаджуються запаси гістонів, тубуліну, ліпофосфопротеїдів та жовтка.
Кортикальний шар ооплазми розміщений у периферійній частині овоцитів. він не має жовтка, містить більшість мітохондрій та гранул глікозаміногліканів. Кортикальний шар відіграє важливу роль в заплідненні (рис. 21).
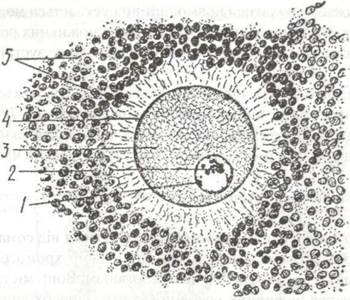
Рис. 21. Яйцеклітина ссавця:
1-ядро; 2-ядерце; 3-цитоплазма; 4-первинна оболонка: 5-вторинна оболонка (фолікулярні клітини).
Яйцеклітини мають дві або три оболонки:
1) первинна оболонка (жовткова або відтелінова, це власне плазмолема)
2) вторинна оболонка - утворена фолікулярними клітинами та їх дохідними. Останні формують блискучу (прозору) зону (від лат. zona реllucsida). Відростки фолікулярних клітин пронизують блискучу зону утворюючи променевий вінець (від лат. corona radiata). Вторинна оболонка виконує трофічну і захисну функції та запобігає поліспермії.
3) третинна оболонка - існує в яйцеклітинах плазунів, яйцекладних, ссавців, та деяких риб (селахій) вона утворюється секретом яйцепроводів.
Для овоцитів характерна полярність, котра пов’язана з кількістю жовтка та розміщенням ядра. Та частина яйцеклітини, д якій накопичу» ться жовток, є вегетативним полюсом, а протилежна, де знаходиться ядро і менше жовтка називається анімальним полюсом.
В залежності від кількості жовтка яйцеклітини поділяються на три групи: оліголецитальні (від гр. oligos - мало, lecithos — жовток), мезолецитальні (від гр. mesos — середній) та полілецитальні (від гр.poli — багато). Залежно від розміщення жовтка яйцеклітини бувають телолецитальні (від гр. telos — кінець) та ізолецитальні (від гр. isos — рівний).
Оліголецитальні овоцити характерні для плацентарних ссавців в зв'язку з внутрішньоутробним розвитком і живленням за рахунок материнського організму. Винятком є овоцити представників примітивних ссавців (клоачні, частково сумчасті), які зберегли основні ознаки своїх попередників — плазунів, у тому числі й телолецитальні овоцити. У телолецитальних овоцитів-жовток концентрується на одному полюсі.
У сільськогосподарських тварин діаметр овоцита коливається від 100 до 140 мкм. Жовтка у них занадто мато, проте в ооплазмі речовини розподілені нерівномірно. На одному боці яйцеклітини нагромаджуються РНП (рибонуклеопротеїди), тому на цій частині поверхні овоцита можна помітити серпоподібної форми скупчення базофільної речовини. Вважають, що цей базофільний серп відповідає і майбутній спині зародка. На майбутньому нейтральному боці його нагромаджується жовток. Таким чином, у овоцитах ссавців ще до запліднення можна спостерігати полярність розміщення поживних речовин.
Овоцити ссавців містять органели загального призначення: ендоплазматичну сітку, мітохондріїт комплекс Гольджі. На периферії цитоплазми знаходиться кортикальний шар, в якому знаходяться мітохондрії та гранули глікозаміногліканів. Включення ооплазми представлені жовтком у вигляді гранул, утворених фосфоліпідами, протеїнами та вуглеводами,. Зовні від кортикального шару знаходиться плазмолема овоцита. Фолікулярні клітини формують і променевий вінець — corona radiata - друга складова вторинної оболонки.
Мезолецитсільні овоцити. Мають розмір до 2-3 мм, розміщені нерівномірно. В ділянці анімального полюса мало жовтка, міститься ядро з великою кількістю ядерець. Більша частина жовтка розміщена з боку вегетативного полюса, на якому жовткові платівки щільно спаковані. Кортикальний шар товщиною 1 мкм містить гранули, а також чорний пігмент. Овоцити амфібії належать до телолецитального типу, чітко полярні, мають сполучнотканинну оболонку, зовні якої знаходиться слизовий шар, що являє собою третинну оболонку.
До полілецитального типу відносять яйцеклітини птахів. Власне яйцеклітиною птахів є жовток, розмір якого у курей досягає 3,5 см. В яйцепроводі яйцеклітина покривається третинними оболонками, до яких належать: зовнішній шар вітелінової мембрани, за її рахунок формуються градинки, які є зв'язками, на яких підвішений жовток, білкова оболонка, дві підшкаралупні оболонки, шкаралупа, побудована із органічних волокон неорганічних речовин. Третинні оболонки виконують трофічну та захисну функції. Полілецитальні та телолецитальні яйцеклітини характерні для деяких риб, плазунів, птахів та яйценесучих ссавців (рис. 22).
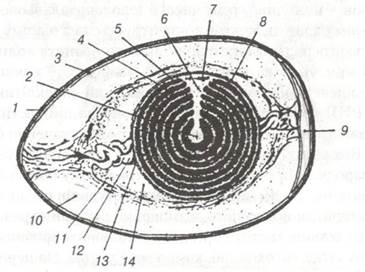
Рис. 22. Схема будови курячого яйця:
1-шкаралупа; 2-латебра; 3-підшкаралупна оболонка; 4-білий жовток; 5-жовтий жовток; 6-ядро Пандера; 7-бластодерма;
8-жовткова оболонка; 9-пові- тряна камера; 10-зовнішній шар білка; 11-волокнистий шар білка; 12-халаза;
13-халазоутворюючий шар; 14-внутрішній шар білка.