ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 2. ЕМБРІОЛОГІЯ
2.3 Гаметогенез
Первинні статеві клітини — гаметобласти виникають у зародка на ранній стадії розвитку. В цей період вони ще не мають статевих особливостей і являють собою великі клітини. Вони відрізняються від інших клітин зародка вмістом глікогену та активними ферментами, серед яких важливе місце належить лужній фосфатазі. За цією ознакою їх виявляють, ставлячи відповідні гістохімічні реакції. Виникають ці клітини у птахів і ссавців в тимчасовому органі — мезодермі жовткового міхура, де формуються первинні кровоносні судини і де в цей період створюються сприятливі умови для живлення. Пізніше первинні статеві клітини переносяться кров'ю або активно переміщуються в зародки гонад — сім'яники або яєчники, де і відбувається їх розвиток (рис.23).
Спосіб поділу статевих клітин, при якому дочірні клітини одержують вдвічі менше хромосом називається мейозом (від лат. meiosis — зменшення). Завдяки цьому поділу забезпечується постійність числа хромосом у наступних поколіннях. В результаті мейозу утворюються статеві клітини з одинарним (гаплоїдним) набором хромосом. Процес мейозу може тривати від кількох днів до кількох років.
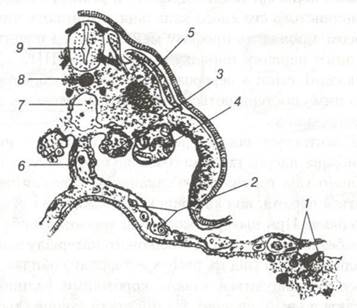
Рис. 23. Схема зрізу через середню частину тіла раннього зародка ссавця, що ілюструє виникнення первинних статевих клітин у ентодермі жовткового міхура і їх міграцію до гонад:
1-первинні статеві клітини; 2-клітини, що мігрують до брижі; 3-клітини, які локалізуються у гонадах; 4-ембріональна нирка;
5-міотом; 6-дорзальна мезодерма; 7-аорта; 8-хорда; 9-нервова трубка.
Розвиток сперматозоїдів - сперматогенез.
Сперматогенез відбувається в звивистих сім’яних канальцях, у яких містяться сперматогенні клітини та клітини Сертолі (сустентоцити), останні утворюють мікросередовище для сперматогенних клітин.
В сперматогенезі розрізняють чотири періоди: розмноження, росту, дозрівання, формування.
На стадії розмноження клітини називаються сперматогоніями. Вони межують з базальною мембраною, тобто займають периферійну частину звивистого сім'яного канальця. Сперматогонії невеликого розміру містять диплоїдний набір хромосом. Серед них існує два різновиди: одні з них мають овальне світле ядро з розсіяним у ньому хроматином. Діляться вони мітозом, збільшуючись кількісно. Це стовбурові клітини. Сперматогонії другого різновиду мають кулясте ядро, що містить зернистий хроматин. Такі сперматогонії перестають ділитися і переходять в стадію росту} перетворюючись у сперматоцити першого порядку. Вони збільшені в розмірі, зміщені в напрямі просвіту звивистого сім'яного канальця, зберігають диплоїдний набір хромосом, проходять профазу мейозу. Перед початком профази у сперматоциті першого порядку подвоюється ДНК, а також відбуваються складні зміни в перерозподілі спадкового матеріалу в ядрі, в зв'язку з цим спостерігаються такі фази: лептонеми, зигонеми, пахінеми, диплонеми.
В стадії лептонеми (від гр. leptos - тонкий, nеmае — нитка) помітна ядерна мембрана, ядерце та хромосоми, що мають вигляд тонких ниток. В зигонемі (від гр. zigotis -з’єднання) — гомологічні хромосоми розміщуються парами, кон’югуючись за довжиною і утворюють біваленти, або діади. При цьому гомологічні хромосоми обмінюються генами, що забезпечує мінливість спадкового матеріалу в ряді поколінь.
На стадії пахінеми (від гр. pachys — твердий) обидві аутосоми спіралізуються, потовщуються, стають коротшими, залишаються у тісному контакті по всій довжині. На цій стадії закінчується кон’югація хромосом.
В диплонемі (ві ф· diplos — подвійний) в кожній із кон'югуючих гомологів з'являються по парі сестринських хроматид (рис. 24). Хроматиди пов'язані центромерою, що перетворює біваленти в тетради. В кожній парі виникає поздовжня щілина, і у двох кон'югованих хромосомах утворюється чотири хроматиди. В тетрадах з'являються перехрести хромосом (хіазми), що свідчить про обмін гомологічними частинами між несестринськими хроматидами в тетраді. В стадії диплонеми зникають сили гомологічного притягання і виникають сили відштовхування між сестринськими хроматидами. Після цього хромосоми ще більше спіралізуються, потовщуються і відокремлюються одна від другої.
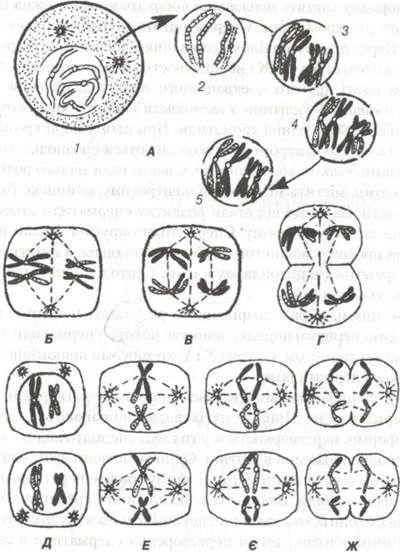
Рис. 24. Схематичне зображення основних стадій мейозу (пояснення в тексті):
А — перше ділення мейозу; 1-лептотена; 2-зиготена; 3-пахітена; 4-диплотена; 5-діакінез.
За періодом росту сперматоцити першого порядку вступають в період дозрівання, який відбувається у два етапи. Після першого — редукційного поділу сперматоцита першого порядку утворюється два сперматоцити другого порядку. Кожний з них одержує половину від кількості хромосом сперматоцита першого порядку. Сперматоцити другого порядку містять диплоїдний набір хромосом. Кожна із хромосом має по дві хроматиди. Сперматоцити другого порядку менших розмірів. Перед другим поділом дозрівання у сперматоциті другого порядку не синтезується ДНК і не подвоюється хромосомний матеріал.
У результаті другого — екваційного поділу дозрівання утворюються сперматиди — клітини з гаплоїдним набором хромосом, кожна з яких складається з однієї хроматиди. При цьому пари хроматид, які з'єднані між собою центромерою, роз'єднуються і розподіляються між сперматидами, стають хромосомами. Сперматиди малого розміру, бідні на хроматин, містять мітохондрії, центросому, комплекс Гольджі та інші органели. Залежно від стадії розвитку сперматиди можуть мати округлу чи витягнуту форму. Сперматиди округлої форми розміщуються біля просвіту звивистого сім'яного канальця, а витягнутої форми — занурюються в цитоплазму клітин Сертолі, в якій вони переходять в період формування.
Таким чином, у фазі дозрівання в результаті мейозу із кожного сперматоцита першого порядку виникає чотири сперматиди з одинарною кількістю хромосом. Статеві X і Y-хромосоми рівномірно розподіляються між сперматидами.
Стадія формування відбувається шляхом перетворення сперматид в сперматозоїди. При цьому їх ядра ущільнюються, набувають овальної форми, перетворюються у голівки сперматозоїдів і займають ексцентричне положення в клітині. Одночасно вони частково занурюються в цитоплазму клітин Сертолі, що зумовлює перетворення сперматид в сперматозоїди. Вважається, що клітини Сертолі забезпечують трофіку та фагоцитують залишки, що виникають в процесі дегенерації неповноцінних клітин. Деталі перетворення сперматиди в сперматозоїда можна виявити за допомогою електронної мікроскопії. Так, частина цитоплазми сперматиди, що містить апарат Гольджі, концентрується на апікальному кінці голівки сперматозоїда. У цій зоні виникає ущільнена гранула — акробласт. Він збільшується в розмірі і у вигляді чохлика покриває ядро. В середині акробласта утворюється акросома. Центросома входить до складу шийки, вона складається із двох центріолей, зміщується в протилежний кінець сперматиди. Проксимальна центріоля прилягає до поверхні ядра, а дистальна поділяється на дві частини. Від краніальної частини дистальної центріолі помітно початок формування джгутика сперматозоїда, каудальна частина центріолі набуває вигляд кільця, зміщуючись по джгутику, це кільце визначає задню межу середньої частини сперматозоїда.
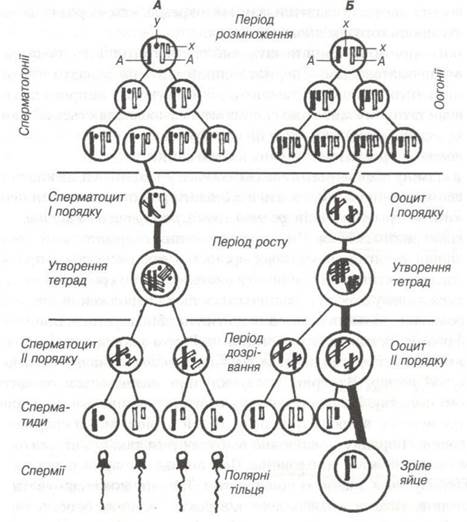
Рис. 25. Схема сперматогенезу (А) і овогенезу (Б).
Батьківські хромосоми чорні, материнські світлі. А — аутосоми; Χ,Υ — статеві хромосоми; в періоді розмноження показані три покоління сперматогоній і оогонії.
Під час росту хвоста цитоплазма сповзає з ядра і концентрується у проміжній частині. Мітохондрії розміщуються спірально між проксимальною центріолею та кільцем. Цитоплазма сперматиди під час її перетворення в сперматозоїд значно редукується. В ділянці голівки
вона зберігається лише у вигляді тонкого шару, що покриває акросому. Незначна її кількість сповзає в дистальному напрямку і утворює тонку оболонку хвостика, а його кінцева частина зовсім не має цитоплазми. Невикористана частина цитоплазми відокремлюється і розпадається у просвіті звивистого сім'яного канальця.
Таким чином, кожний початковий сперматогопій дає початок чотирьом сперматозоїдам. Сперматогенний епітелій занадто чутливий до впливу інтоксикації, авітамінозу, дії іонізуючого випромінювання. При недостатньому живленні сперматогенез послаблюється або зовсім зникає, а сперматогенний епітелій атрофується.
Овогенез — розвиток статевих клітин самки.
На відміну від сперматогенезу, овогенез у тварин різних класів значно різноманітніший і залежить від біології розмноження. Він перебігає протягом трьох періодів: розмноження, росту та дозрівання.
Період розмноження. Якщо розмноження сперматогоній починається після досягнення статевої зрілості і продовжується протягом усієї статевої активності самця, то в овогенезі період розмноження відбувається лише у зародка і закінчується після народження плода. Після народження кількість статевих клітин не збільшується. Вони мають малий розмір, їх називають овогоніями. Разом з покриваючими їх фолікулярними клітинами називаються примордіальними фолікулами.
Період росту. В період росту клітини називаються овоцитами першого порядку. З самого початку свого існування овоцит першого порядку вступає в профазу мейозу. В цей період, як і в сперматоцитах першого порядку, послідовно відбуваються такі стадії: лептонема, зигонема, пахінема й диплонема. Цей період у тварин різних класів характеризується значною повільністю. Так, він може відбуватися за кілька днів, тижнів і навіть десятків років. Останнє спостерігається у ссавців, де розмноження овогоній закінчується іще до народження плода, а овуляція настає не раніше статевої зрілості. Після диплонеми хромосоми розходяться, деспіралізуються, стають непомітними до кінця періоду росту. Овоцит першого порядку інтенсивно синтезує речовини за допомогою хромосом.
Нагромадження поживних речовин овоцитом першого порядку поділяють на пуевітелогенез (фаза невеликого пості А та вітелогенез (фаза великого роасту). Превітологенез характеризується ростом цитоплазми і переважним синтезом інформаційної РНК, яка пізніше буде матрицею для синтезу білків. Такий інтенсивний синтез у овогенезі інформаційної РНК пов'язаний з тим, що він іде на запас. Не більше третини його використовується безпосередньо в овогенезі, тоді як основна частина білкового синтезу починається після запліднення. У цей час в цитоплазмі утворюються органели, необхідні для синтезу: гранулярна цитоплазматична сітка, мітохондрії, комплекс Гольджі.
У фазі великого росту синтезуються запасні поживні речовини, що нагромаджуються у цитоплазмі і використовуються лише після запліднення. Основні з них жовток, жир, глікогени. Жовток являє собою кристалізовані речовини білкової природи у формі фосфопротеїнів. Кристали мають вигляд гранул чи платівок різної форми. Кількість жовтка, що нагромаджується в яйцевій клітині, неоднакова у тварин різних груп. В полілецитальних яйцеклітинах, наприклад курей, жовток відкладається інтенсивно. Так, за добу відкладається шар жовтка товщиною до 2,5 мм. У курей перед овуляцією яйцеклітина досягає 35 мм (Б. Карлсон, 1983). Яйцеклітини ссавців значно менші за розміром. їх ріст може продовжуватися роками, наприклад у корів — 10 років і більше.
В синтезі жовтка в цей час бере участь весь організм, особливо печінка. Запас поживних речовин у вигляді жовтка називають дейтоплазмою. В процесі нагромадження жовтка майже завжди виявляють полярність овоцита. Полюс, багатий на жовток, називають вегетативним, протилежний, куди зміщується ядро — анімальним. У фазі великого росту органели розсіюються по цитоплазмі з подальшою концентрацією на периферії.
Період дозрівання відбувається після проникнення в яйцеву клітину сперматозоїда (у аскариди), у ланцетника спермій проникає після першого поділу; у більшості тварин він проникає в яйце лише після дозрівання, яке настає після виходу яйцеклітини із яєчника.
Дозрівання овоцита - це складні перетворення у ядрі. Як і при сперматогенезі, воно складається із двох послідовних поділів, які надають яйцеклітині гаплоїдного стану. Однак, в дозріванні яйцеклітини, на відміну від сперматогенезу, один первинний овоцит утворює лише одну зрілу яйцеклітину, яка повністю зберігає жовток, що утворився в період росту, і три неповноцінні клітини — полярні (редукційні) тільця. Вони містять лише ядро без цитоплазми. При цьому ядро первинного овоцита ділиться порівну між двома дочірніми клітинами, тобто між овоцитом другого порядку і першим редукційним тільцем. Подальший поділ приводить до того, що овоцит другого порядку також ділиться нерівномірно — виникає велика зріла яйцеклітина та друге редукційне тільце. В подальшому редукційні тільця гинуть. їх призначення полягає у тому, що дозріваюче за їх допомогою яйце звільняється від половини хромосом. Під час другого поділу дозрівання яйцеклітина втрачає свою центросому.