ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 3. ЗАГАЛЬНА ГІСТОЛОГІЯ
3.5 Нервова тканина
3.5.5 Нервові закінчення
Термінальні апарати нервових волокон — нервові закінчення за їх функціональним значенням поділяють на три види: рецептори, ефектори та міжнейронні синапси.
Рецептори (чутливі нервові закінчення) — спеціалізовані термінальні структури дендритів нейронів, пристосовані до сприйняття подразнень, що надходять до організму. Залежно від локалізації і специфічності нервової регуляції життєдіяльності організму рецептори поділяють на екстерорецептори (сприйняття подразнень із зовнішнього середовища) та інтерорецептори (подразнення власних тканин організму). Різновидом інтерорецепторів є пропріорецептори — нервові закінчення у м'язах та сухожиллях, які беруть участь у регуляції рухів і положенні тіла у просторі. Залежно від природи подразнень, останні поділяють на термо-, механо-, баро-, хемо-, но- ціорецептори та інші.
Чутливі нервові закінчення надзвичайно різноманітні за структурною організацією. Вони поділяються на вільні нервові закінчення, які складаються лише із термінальних розгалужень осьового циліндра дендрита чутливого нейрона та невільні — крім осьового циліндра, містять також клітини нейроглії. Якщо невільні рецептори оточені сполучнотканинною капсулою, їх називають капсульованими; ті невільні рецептори, що не мають сполучнотканинної капсули, мають назву некапсульовані. Рецепторні закінчення або рецепторно-нейральні синапси складаються із спеціалізованих клітин, здатних реагувати на подразник і викликати деполяризацію плазмолеми зв'язаного з ним дендрита аферентного нейрона.
Прикладом вільного рецептора можуть бути деревоподібні та кущеподібні нервові закінчення епітеліальної тканини. При формуванні таких закінчень, мієлінові нервові волокна, підходячи до епітеліального пласта, втрачають мієлінову оболонку, а їхні осьові циліндри утворюють кінцеві розгалуження, що розміщуються між окремими епітеліоцитами. За функцією це, як правило, термо- та ноціорецептори. Вільні нервові закінчення можуть обплітати у вигляді кошика волосяні фолікули, реєструючи зміщення у просторі окремих волосин, відіграючи роль механорецепторів.
До складу рецепторного апарату багатошарових епітеліїв належать поодинокі чутливі епітеліальні клітини, так звані дотикові епітеліоцити Меркеля. Ці електронно-прозорі клітини мають сплющені ядра та осміофільні гранули. До їх базальної частини прилягають чутливі нервові закінчення у вигляді дисків (дотикові меніски Меркеля). Нейроепітеліальні клітини Меркеля мають постійне місце в епітеліальному шарі, незважаючи на постійний рух еиітеліоцитів від базального до дистального полюса. Клітини Меркеля — типовий приклад нейроепітелію, епітеліоцити якого пристосувалися до рецепції, вони виконують функцію механорецепції.
Надзвичайно різноманітні рецепторні апарати сполучної тканини представлені двома групами невільних нервових закінчень. У капсульованих рецепторах нервові терміналі аферентних волокон, як правило, оточені нейролемоцитами і допоміжними елементами сполучнотканинного походження. До капсульованих чутливих закінчень, залежно від будови, належать: тільця Фатер-Пачіні, Гольджі-Маццоні, Мейснера, кінцеві колби Краузе (генітальні тільця).
Найбільш поширені рецептори сполучної тканини — це пластинчасті тільця Фатер-Пачіні (рис. 79). У їх складі розрізняють внутрішню колбу та зовнішню капсулу. Навколо розгалужень осьового циліндра (термінальний відділ чутливого нервового волокна), яке втратило мієлінову оболонку, скупчуються видозмінені нейролемоцити, що і утворюють внутрішню колбу. Капсула тілець Фатер-Пачіні складається із великої кількості сполучнотканинних пластинок, утворених фібробластами та спірально орієнтованими пучками колагенових волокон. На межі внутрішньої колби та зовнішньої капсули розміщуються клітини, які, очевидно, визначаються як гліальні. Ці гліоцити утворюють синапси із розгалуженням осьового циліндра. Є припущення, що нервовий імпульс генерується в умовах зміщення зовнішньої капсули щодо внутрішньої колби.
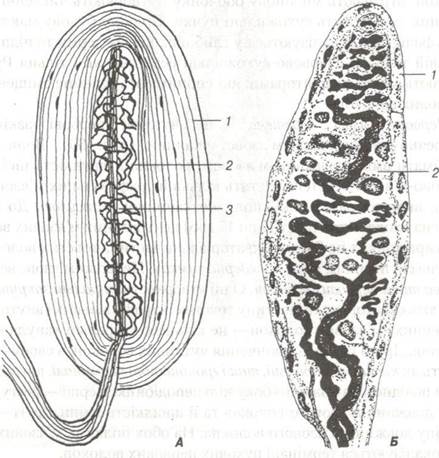
Рис. 79. Схема будови капсульованих рецепторів:
А — тільце Фатер-Пачіні; Б — дотикове тільце Мейснера; 1-зовнішня капсула (сполучнотканинні клітини);
2-внутрішня колба (нейрогліальні клітини); 3-осьовий циліндр.
Тільця Гольджі-Маццоні менші від тілець Фатер-Пачіні, мають тонку капсулу і, порівняно, велику внутрішню колбу. Ці структури виконують функцію барорецепції у шкірі, серозних та слизових оболонках.
Тільця Мейснера (дотикові) — це рецептори тактильної чутливості, які локалізуються у сосочковому шарі дерми. їх олігодендроцити орієнтовані перпендикулярно до поверхні шкіри і мієлінові волокна, втрачаючи мієлінову оболонку, контактують із поверхнею гліальних клітин колби. Колагенові волокна сполучнотканинної капсули можуть заповнювати простори між нейролемоцитами і терміналями чутливого нервового волокна.
Кінцеві колби Краузе є механо- і терморецепторами кон'юнктиви, язика, а видозмінені структури (генітальні тільця) — зовнішніх статевих органів. Характерною особливістю будови колб Краузе є дуже тонка сполучнотканинна капсула. Мієлінове волокно, входячи в капсулу, втрачає мієлінову оболонку і закінчується колбоподібним розширенням. Генітальні тільця мають подібну будову, за винятком того, що в капсулу проникає не один осьовий циліндр, а декілька, які, розгалужуючись, утворюють сіточку безмієлінових волокон.
Нервово-сухожильні веретена (сухожильні органи Гольджі) утворені товстими мієліновими волокнами, які, підходячи до колагенових волокон, втрачають мієлінову оболонку і утворюють численні розгалуження, обплітають сухожильні пучки. Подібну будову мають тільця Руффіні, що локалізуються у глибоких шарах дерми та підшкірній жировій тканині. Нервово-сухожильні веретена, як і тільця Руффіні вважають механорецепторами, які сприймають взаємне зміщення колагенових волокон.
Нервово-м'язові веретена— це чутливі нервові закінчення поперечно-посмугованої м'язової тканини (рис. 80). Вони здатні сприймати зміну довжини м'язового волокна і швидкість цієї зміни. Нервово-м'язові веретена містять кілька інтрафузальних м'язових волокон, вкритих загальною сполучнотканинною капсулою. До складу веретена входять два товстих по 17 мкм центральних м'язових волокна та чотири тонких по 8 мкм. Екваторіальна частина товстих волокон заповнена скупченням ядер — «ядерна сумка». Нервово-м'язове веретено має два типи нервових волокон. Одні утворюють спіральні закрутки, що оточують екваторіальну частину товстих інтрафузальних (внутрішньо- веретенних) м'язових волокон — це кільцево-спіральні «анулярні» закінчення. Друга група — закінчення чутливих волокон, в своєму складі містить як кільцево-спіральні, так і гроноподібні терміналі, що розміщуються по одному з кожного боку кільцеподібних. Перші — реагують на зміну довжини м'язового волокна та її швидкість зміни, другі — лише на зміну довжини м'язового волокна. На обох полюсах м'язових волокон локалізуються терміналі рухових нервових волокон.
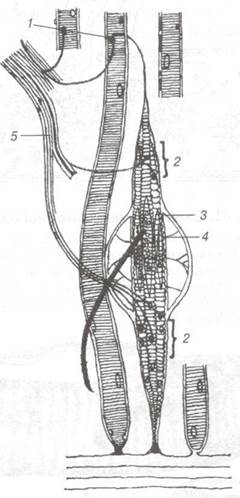
Рис. 80. Схема будови нервово-м'язового веретена:
1-моторні бляшки екстрафузальних м'язових волокон; 2-моторні бляшки інтрафузальних м'язових волокон; 3, 4-ядерна сумка (скупчення ядер); 5-м'язовий нерв.
Ефектори — структури, утворені закінченням аксонів нейроцитів, які бувають рухові та секреторні. Нервово-м'язові закінчення у скелетних м'язах утворені терміналями аксонів нейроцитів, рухових ядер спинного та моторних — головного мозку. При наближенні до м'язового волокна мієлінове волокно втрачає мієлінову оболонку, його осьовий циліндр розгалужується і разом із саркоплазмою міосимпласта занурюється у м'язове волокно (рис. 81).
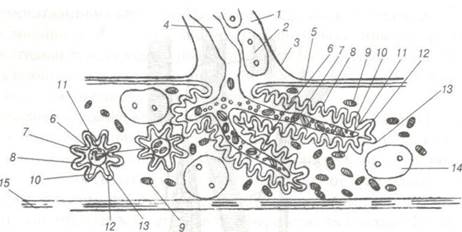
Рис. 81. Схема будови моторної бляшки:
1-цитоплазма нейролемоцита; 2-ядро; 3-нейролема; 4-осьовий циліндр; 5-сарколема; б-кінцеві гілочки нервового волокна в повздовжньому та поперечному розрізах; 7-мітохондрії аксоплазми; 8-первинний синаптичний простір; 9-саркосоми;
10-вторинний синаптичний простір; 11-синаптичні пухирці; 12-пресинаптична мембрана; 13-постсинаптична мембрана;
14-ядро моторної бляшанки; 15-міофібрила.
У цій ділянці утворюється аксом'язовий синапс, де аксолема відіграє роль пресинаптичної мембрани, а сарколема м'язового волокна є іюстсинаптичною мембраною (рис. 82). Медіатор цих синапсів — ацетилхолін, який при збудженні вивільняється і через пресинаптичну мембрану проникає у синаптичну щілину. На поверхні постсинаптичної мембрани ацетилхолін зв'язується із холінорецепторами і стає хімічним сигналом для збудження м'язового волокна.
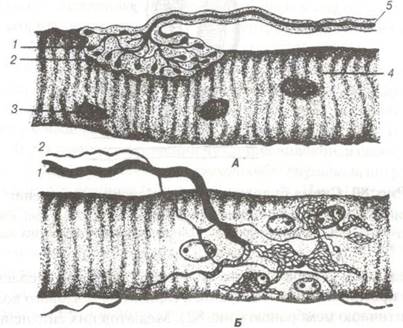
Рис. 82. Ефекторне нервове закінчення (моторна бляшка):
А — вигляд у профіль: 1,2 — терміначі мієлінового нервового волокна; 3-ядра м'язових волокон; 4-м'язове волокно; 5-мієлінове волокно; Б — вигляд зверху: 1- мієлінове волокно; 2- безмієлінове волокно; 3-ультратермінальне волокно (те, що входить до моторної бляшки і закінчується на іншій).
Еферентні закінчення у гладенькій м'язовій тканині мають простішу будову. Окремі нервові закінчення на поверхні гладеньких міоцитів утворюють варикозні терміналі. Медіатор цих закінчень — ацетилхолін, або адреналін.