ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 4. СПЕЦІАЛЬНА ГІСТОЛОГІЯ
4.2 Органи чуття
4.2.2 Орган слуху та рівноваги
Органом рівноваги та слуху є вухо. Природа сприйняття цих різних відчуттів подібна, оскільки зміна положення тіла в просторі чи звукові коливання викликають переміщення рідини всередині цих органів, що, в свою чергу, викликає подразнення війок чутливих клітин, що і породжує відчуття.
Вухо — периферійна частина слухового та вестибулярного аналізаторів. Це складний комплекс структурних елементів, в яких розміщуються рецептори, здатні забезпечити сприйняття звукових вібраційних та гравітаційних сигналів.
Вухо складається з трьох частин: зовнішнього, середнього та внутрішнього вуха (рис. 96).
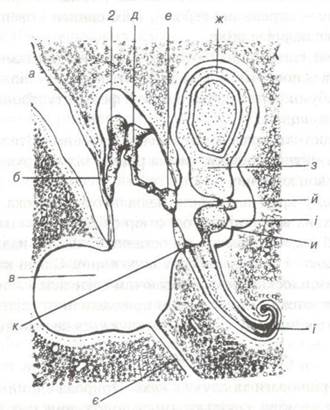
Рис. 96. Схема будови вуха.
a-зовнішній слуховий прохід; б-барабанна перетинка; в-барабанна порожнина; г-молоточок; д-ковадло; е-стременце; є-слухова труба; к-напівокру- глий канал; з-маточка; и-мішечок; і-переддвір’я; ї-завитка; й-овальне і к- кругле вікна.
До зовнішнього вуха відносять вушну раковину, зовнішній слуховий прохід, який закінчується барабанною перетинкою, що відмежовує зовнішнє вухо від середнього.
Середнє вухо являє собою барабанну порожнину кам'янистої кістки із слуховими кісточками: молоточком, ковадлом і стременем. За допомогою слухової (євстахієвої) трубки середнє вухо з'єднується з глоткою.
Внутрішнє вухо поділяється на кістковий і перетинчастий лабіринти. Кістковий лабіринт — це система порожнин і каналів всередині скелястої частини кам'янистої кістки. Внутрішнє вухо відмежовується від середнього овальним і круглим віконними — отворами в перетинці між барабанною і скелястою частинами кістки, закритими сполучнотканинними мембранами.
В кістковому лабіринті розрізняють три відділи: кісткову завитку, три кісткових напівокруглих канали, які розміщені у трьох взаємопер- пендикулярних площинах, і переддвір'я. В середині кісткового лабіринту знаходиться перетинчастий лабіринт, до складу якого входить орган рівноваги з перетинчастими напівкруглими каналами, овальним та круглим мішечками і орган слуху у вигляді перетинчастої завитки. Між стінками кісткового та перетинчастого лабіринтів залишаються простори, заповнені рідиною — перилімфою.
Порожнини перетинчастого лабіринту також містять рідину — ендолічфу. Ці порожнини з'єднуються через щілини в кам'янистій кістці з підоболонковими просторами головного мозку.
Розвиток вуха починається з випинання ектодерми в ділянці заднього мозкового міхура у вигляді слухової ямки, яка незабаром від- шнуровується і перетворюється у слуховий міхурець (рис. 97). Не- йроепітелій слухового міхурця диференціюється на слухові (чутливі) і опорні клітини. Слухові клітини на своїй вільній поверхні мають війки, що зв’язані з відростками біполярних клітин спинального ганглію, закладеного поряд. В одній частині міхурця слухові клітини розміщуються острівцями, в другій — смужками. Ці дві частини поділяються і перша згодом стає зачатком органа рівноваги, а друга — органа слуху. У процесі подальшого розвитку з'являються плоскі випинання, з яких утворюються півкруглі канали.
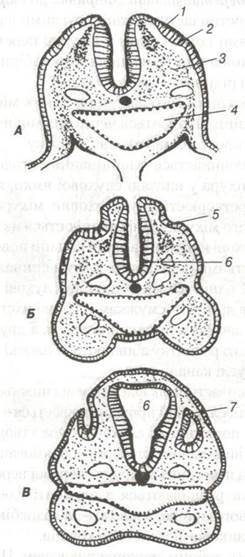
Рис. 97. Схема розвитку слухового пухирця ембріона людини (за Ареєм):
А — стадія 9 сомітів; Б — стадія 16 сомітів; В — стадія 30 сомітів;
2-слухова плакода; 3-мезодерма; 4-глотка; 5-слухова ямка; 6-мозковий пухирець; 7-слуховий пухирець.
Орган слуху розвивається, як відросток від нижньої частини слухового міхурця. Зовнішній слуховий прохід утворюється з першого зябрового кармана. Барабанна порожнина і слухова труба утворюються з першого глоткового кармана випинання ектодерми, спрямованого в бік зябрового кармана, а барабанна перетинка з першої зябрової перетинки.
Слухові кісточки розвиваються з елементів зябрових дуг: молоточок — з меккелевого хряща, ковадло — з піднебінно-квадратного, а стремінце — з під'язиково-щелепового хрящів.
Вушна раковина є добрим звукоуловлювачем. Ця функція особливо розвинена в деяких тварин, у яких рефлекторно-вушна раковина полегшує знаходження джерела звуку. Вушна раковина вкрита шкірою і волоссям. У шкірі закладені сальні і потові залози. Основа вушної раковини складається з еластичного хряща. До неї прикріплені м'язи з поперечнопосмугованої м’язової тканини.
Зовнішній слуховий прохід вистелений продовженням шкіри вушної раковини, в якій містяться альвеолярні сальні та видозмінені трубчасті потові залози, покриті одношаровим циліндричним епітелієм і продукують речовину, що містить слиз та пігменти (вушну сірку).
Барабанна перетинка складається з щільної сполучної тканини, з боку зовнішнього слухового проходу покрита багатошаровим плоским епітелієм, з боку барабанної порожнини — одношаровим, а по краях — миготливим. В сполучній тканині знаходиться ручка молоточка.
Середнє вухо. Барабанна порожнина вистелена слизовою оболонкою у вигляді тонкого, зрощеного з надкісницею (окістям) сполучнотканинного шару, покритого двошаровим кубічним, а місцями — призматичним епітелієм. Ця оболонка покриває також слухові кісточки. Під епітелієм залягає густа сітка капілярів, а також нервове сплетення з гангліозними клітинами.
Слухові кісточки передають звукові коливання від барабанної перетинки до овального віконця і перилімфи. Вони побудовані із пластинчастої кісткової тканини, а на суглобових поверхнях містяться залишки хряща. М’язи їх є поперечнопосмугованою м’язовою тканиною. За допомогою останніх збільшується сила звукових коливань при натягуванні барабанної перетинки або орган слуху захищається від надмірного подразнення, розтягуючи ланцюжок слухових кісточок.
Слухова (євстахієва) труба з'єднує порожнину середнього вуха з глоткою і через неї з навколишнім середовищем, зрівноважуючи тиск повітря на барабанну перетинку. Біля барабанної порожнини є кісткова стінка, а в глотковому кінці — хрящова. Слизова оболонка покрита багаторядним миготливим епітелієм і містить лімфатичні вузлики й слизові залози. В стінці евстахієвої труби проходить багато кровоносних судин і нервів з невеличкими гангліями.
Внутрішнє вухо складається з кісткового й перетинчастого лабіринтів. У кістковому лабіринті є три частини: переддвір'я, три півколові канали і завиток, всередині яких знаходяться відповідні перетинчасті частини. Перетинчаста частина переддвір'я представлена двома мішечками — утрикулюсом і сакулюсом. Утрикулюс сполучається з перетинчастими півколовими каналами, розміщеними у трьох взаємно перпендикулярних площинах. Один кінець кожного каналу колбоподібно розширений і називається ампулою. Сакулюс (від лат. sacculus — мішечок) через невелику протоку з'єднується з перетинчастим каналом завитки. В стінці кожного мішечка є підвищення, які називають плямами або макулами (від лат. macula — пляма), а в стінці ампул — гребінцями.
Макули (рис. 98А), плями утрикулюса(маточки), сакулюса та гребінці — крісти ампул напівкругових каналів є тими чутливими пристосуваннями, в яких виникають сигнали при зміні положення голови або тіла в просторі. Ці спеціалізовані ділянки вестибулярного апарату покриті епітелієм, в якому містяться два види клітин: рецепторні (волоскові ) і опорні.
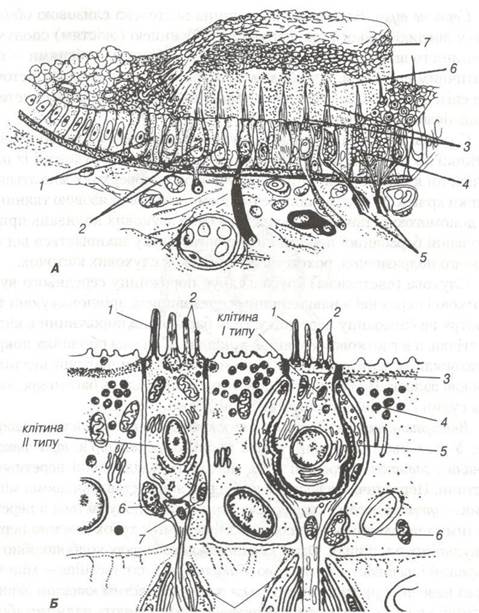
Рис. 98. Манула.
А-будова на світлооптичному рівні (схема): 1-підтримуючі епітеліоцити; 2- сенсорні епітеліоцити (за Кольмером); 3-волоски;
4-нервові закінчення; 5- мієлінові нервові волокна; 6-студениста отолітова мембрана; 7-отоліти.
Б — будова на ультрамікроскопічному рівні (схема): 1-кіноцилія; 2-стереоцилії; 3- кутикула; 4-підтримуючі епітеліоцити;
5-чашоподібне нервове закінчення; 6- еферентне нервове закінчення; 7-аферентне нервове закінчення;
8-мієлінові нервові волокна(дендрити).
Високі опорні клітини своїми розширеними основами розміщуються на базальній мембрані. На їх апікальному полюсі, що досягає вільної поверхні епітеліального пласта, розвинені мікроворсинки.
Між опорними клітинами, правильно чергуючись, розміщуються волоскові чутливі клітини, які своїми базальними поверхнями не торкаються базальної пластинки. Основи цих клітин контактують з аферентними і еферентними нервовими закінченнями, а на апікальній поверхні їх розміщується від 35 до 95 волосків (цілій). Серед волосків один рухомий і найдовший — кіноцилій, інші — нерухомі і по висоті розміщуються східчасто — стереоцилгї. Рецепторні клітини поділяються на два типи. Клітини першого типу мають форму колбочок з закругленими основами, занурені в чашоподібне розширене закінчення аферентного нервового волокна, з яким утворюють синаптичні контакти. Клітини другого типу циліндричної форми і до їх основи примикають аферентні та еферентні нервові закінчення, що утворюють характерні синапси (рис. 98Б).
Поверхня епітелію макули покрита желатиноподібною масою — отолітовою мембраною, в якій вміщені кристали кальцитів — отоліти та статоконії. При переміщенні тіла в просторі отолітова мембрана, зміщуючись, згинає волоски рецепторних клітин, що викликає їх збудження або гальмування. Відхилення волосків від стереоцилій та кіноцилію викликає збуджуючий ефект, а від кіноцилію до стереоцилій — гальмівний. В епітелії макули різнополярні волоскові клітини розміщуються групами, у результаті чого під час ковзання отолітової мембрани в один бік стимулюється лише певна група клітин, яка регулює тонус певних м'язів тулуба.
Поверхня епітелію гребінців покрита желатиноподібною речовиною за формою куполу висотою до 1 мм, який може закривати просвіт ампули. В речовині куполу знаходяться волоски рецепторних клітин, субмікроскопічна будова яких і їх іннервація подібні з клітинами макули. Подразником для рецепторних клітин гребінців напівкругових каналів є кутове прискорення голови в площині даного каналу. При повороті голови у напівкруговому каналі починає рухатися ендолімфа, викликаючи зміщення купола й згинаючи волоски рецепторних клітин, що призводить до збільшення чи зменшення частоти імпульсацій цих клітин, яка передається нервовим закінченням, що підходять до волоскових клітин. На відміну від рецепторів напівкругових каналів, що реагують на кутові прискорення, отолітові рецептори утрикулюса і сакулюса реагують на лінійні прискорення.
Аналізатор рівноваги. Збудження від рецепторних клітин вестибулярного апарату поширюється по дендритах біполярних клітин, тіла яких розміщуються в вестибулярному ганглії. Аксони цих клітин у складі волокон вестибулярного нерва йдуть до нервових клітин вестибулярних ядер довгастого мозку свого боку. Комплекс вестибулярних ядер довгастого мозку є першим пунктом, де відбувається первинна оцінка інформації про рух та положення тіла і голови в просторі.
Від клітин вестибулярних ядер нервові волокна спрямовуються до нейронів зорового горба, які за допомогою аксонів зв'язані з нервовими клітинами високої ділянки кори великих півкуль. Таким чином, шлях від рецепторного апарату органу рівноваги до кори півкуль охоплює три нейрони: біполярні клітини вестибулярного ганглію, нейрони вестибулярних ядер і клітини зорового горба.
Завитка — частина внутрішнього вуха, де розміщуються рецептори, що сприймають звукові коливання. Завитка у вигляді кісткового спірального каналу всередині кам'янистої кістки закручена у вигляді раковини навколо своєї осі і утворює у тварин до п 'яти витків. Частини завитки, що спрямовані до осі, називають внутрішніми, а в протилежний бік — зовнішніми. По всій довжині завитки на внутрішній частині стінки каналу розміщується кістковий виступ — спіральна пластинка з потовщеним періостом — спіральним лімбом (від лат. limbus — край). Останній ділиться на дві губи: верхню — вестибулярну та нижню — барабанну. Заглиблення між ними називають спіральним жолобком. В основі спіральної пластинки розміщується спіральний ганглій. Потовщення окістя на зовнішній поверхні стінки кісткового завиткового каналу назване спіральною зв'язкою.
Між спіральною пластинкою і спіральною зв'язкою натягнуті дві сполучнотканинні перетинки, які у вигляді спіралі тягнуться вздовж усього завиткового каналу.
Одна з них — базилярна мембрана з внутрішнього боку переходить у барабанну губу лімба.
Друга — вестибулярна мембрана одним боком з'єднана з вестибулярною губою, а другим — з спіральною зв'язкою на деякій відстані від місця кріплення базилярної мембрани. В основі базилярної мембрани знаходяться тонкі колагенові волокна, довші на вершині завитки і короткі в її основі. Між волокнами і фібрилами міститься основна гомогенна речовина, до складу якої входять глікозаміноглікани.
Таким чином, по всій довжині, аж до вершини завитки, кістковий канал поділений двома перетинками на три канали або драбини.
Верхній канал — вестибулярна драбина починається від овального віконця і продовжується до вершини завитки.
Нижній канал — барабанна драбина починається від круглого віконця, а на вершині, в місці з'єднання вестибулярної і базилярної мембран, через вузький отвір — гелікотрему з'єднується з вестибулярною драбиною. Простір обох драбин заповнений перилімфою.
Середня драбина або перетинчастий лабіринт завитки не з'єднаний з порожнинами інших каналів і заповнений ендолімфою. На поперечному розрізі завитковий канал має форму трикутника (рис. 99), сторони якого утворені вестибулярною і базилярною мембранами та судинною смужкою, яка лежить на зовнішній стінці кісткової завитки.
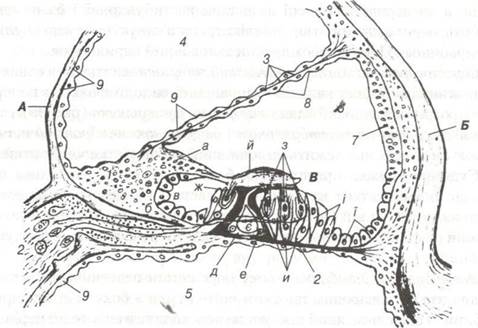
Рис. 99. Схема будови частини завитки на поперечному зрізі.
А — спіральна пластинка; 1- лімб; а-вестибулярна губа; б-барабанна губа; в-спіральний жолоб; г-спіральний ганглій; Б- спіральна зв'язка; 2-базальна мембрана; 3-вестибулярна мембрана; 4-вестибулярна драбина; 5-барабанна драбина; 6-перетинчастий канал завитки; 7-еудинна смужка; 8-одношаровий плоский епітелій; 9-ендотелій; В — спіральний (кортієв) орган; д — внутрішня стовпчаста клітина; е- зовнішня стовпчаста клітина; є- тунель; ж — внутрішні волоскові клітини; з -зовнішні волоскові клітини;
и — зовнішні фалангові клітини; і — зовнішні пограничні клітини; ї — зовнішні підтримуючі клітини; й — покривна текторіальна пластинка
Судинна смужка представлена багаторядним епітеліальним шаром, що розміщується на базальній пластинці. Серед високих епітеліальних клітин у ній багато кровоносних капілярів. Вважається, що епітелій судинної пластинки виконує секреторну функцію — продукує ендолімфу.
Вестибулярна мембрана з боку порожнини перетинчастого каналу покрита одношаровим плоским епітелієм, а з боку вестибулярної драбини — епітелієм, який поступово переходить в ендотелій періосту. Базилярна пластинка з боку барабанної драбини також покрита тонким шаром ендотелію, під яким зустрічаються кровоносні капіляри. З боку порожнини середнього, тобто перетинчастого каналу завитки, на базальній пластинці розміщується спеціалізований епітелій, що утворює звукосприймаючий апарат слухового аналізатора — спіральний (кортієв) орган.
Кортієв орган складається із внутрішніх і зовнішніх клітин двох типів (рис. 100): рецепторних (волоскових) і підтримуючих (опорних). Останні своїми основами розміщуються на базилярній пластинці, що знаходиться між комплексом епітеліальних клітин спірального органу і сполучнотканинною частиною базилярної пластинки.
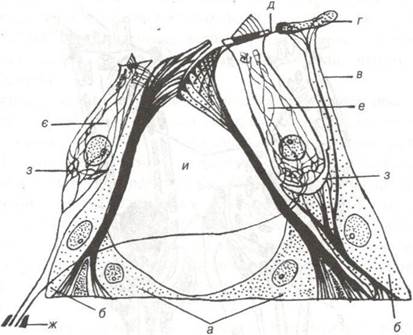
Рис. 100. Рецепторні та підтримуючі клітини спірального органа (за Кольмером):
а-клітини-стовпи; б -фалангові клітини; в-фаланговий відросток; г-фаланго- ва пластинка; д-сітчаста мембрана; е-зовнішня та є — внутрішня волоскові клітини; ж-нервові волокна, що формують синапси на слухових клітинах; з- синапси на слухових клітинах;
и-спіральний тунель.
Розрізняють кілька різновидів опорних клітин: стовпи та фалангові. Клітини-стовпи по довжині спірального органу розміщуються в два ряди (внутрішні і зовнішні стовпи). Розширена основа цих клітин контактує з базилярною пластинкою, апікальними полюсами клітини — стовпи нахилені косо один до одного й утворюють своєрідне склепіння, що прикриває трикутний канал — тунель, заповнений ендолімфою. По тунелю проходять безмієлінові нервові волокна, в яких містяться дендрити нейронів спірального ганглію. Цитоплазма стовпів дуже пружна (еластична) завдяки наявності в ній великої кількості тонофібрил.
В безпосередній близькості від зовнішніх клітин-стовпів розміщується три ряди зовнішніх фалангових клітин. Це циліндричної форми клітини, на апікальному полюсі яких є чашовидне впинання і фаланговий відросток, що доходить до поверхні спірального органа і закінчується пластинкою. Фалангові пластинки, з'єднуючись одна з одною, утворюють сітчасту мембрану, в отворах якої розміщуються верхні кінці слухових клітин, а тіло їх прилягає до внутрішнього боку фалангового відростка.
Таким чином, рецепторні клітини відокремлені одна від одної відростками фалангових клітин. У цитоплазмі останніх по всій осі клітини проходить пучок тонофібрил, які продовжуються у відросток. Зовні від фалангових клітин розміщуються пограничні клітини, на апікальній поверхні яких, є велика кількість мікроворсинок, а в цитоплазмі краплі ліпідів, вакуолі, глікоген, що свідчить про їх трофічну функцію.
Поступово, зменшуючись у висоті, пограничні зовнішні клітини переходять у низькі підтримуючі клітини, які вкривають останню частину базилярної мембрани і переходять в епітелій судинної смужки. Із внутрішнього боку — один ряд фалангових клітин і далі внутрішні пограничні циліндричні клітини, зменшуючись у висоті, переходять у кубічний епітелій спірального жолоба.
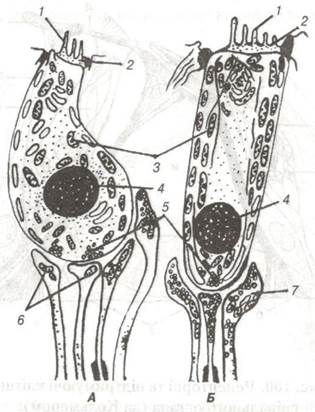
Рис. 101. Ультраструктурна організація внутрішньої (А) та зовнішньої (Б) слухових волоскових клітин (схема):
1-волоски; 2-кутикула; 3-мітохондрії; 4-ядра: 5-синаптичні пухирці в рецепторних клітинах; 6-світлі та 7 — темні нервові закінчення.
Рецепторні — волоскові клітини (рис. 101) знаходяться по обидва боки від клітин-стовпів, причому так, що внутрішні волоскові клітини розміщуються в один, а зовнішні в три ряди. По всій довжині спірального органу налічується до 25 тис. рецепторних клітин, які своїми закругленими основами прилягають до заглиблення на апікальній поверхні фалангових клітин. Таким чином, слухові клітини не мають безпосереднього контакту з базальною пластинкою, а ядра цих клітин розміщені на базальному полюсі. В цитоплазмі таких клітин міститься значна кількість мітохондрій, ендоплазматичної сітки та глікогену.
На апікальній поверхні рецепторних клітин розміщена кутикулярна пластинка з слуховими волосками - стереоциліями (від гр. stereos - об’ємний, від лат cilium — війка). Електронномікроскопічними методами встановлено, що на внутрішніх рецепторних клітинах у вигляді прямолінійної щіточки розміщується 40-50 коротеньких волосків, а на зовнішніх — до 120 довших волосків, що мають вигляд зігнутої (V-noдібної) щіточки.
На вершині волоскових клітин є стрічкоподібна пластинка желеподібної консистенції — покривна мембрана, до складу якої входить прозора основна речовина з вмістом глікозаміногліканів та тоненьких тонофібрил. Одним краєм покривна мембрана з'єднується з верхнім боком вестибулярної губи спірального лімба, а другим, що має на поперечному розрізі форму язичка, по всій довжині контактує з волосковими клітинами, волоски яких занурені в основну речовину покривної мембрани.
Під час звукового збудження, коливання барабанної перетинки через систему слухових кісточок середнього вуха викликає коливання мембрани овального віконця і перилімфи вестибулярної і барабанної драбини.
Коливання перилімфи передаються на вестибулярну мембрану, а потім на порожнину перетинчастого каналу завитки, спричинюючи рух ендолімфи та базилярної мембрани. Експериментально доведено, що кожній висоті звуку відповідає певна довжина базилярної мембрани, охопленої коливальним процесом.
При дії на вухо звуків низької частоти базилярна мембрана зміщується по всій довжині — від основи до вершини завитки. При цьому зміщуються волоски стосовно покривної (текторіальної) мембрани і збуджуються рецепторні клітини.
При дії звуків високої частоти в коливальний процес залучається базилярна мембрана лише на обмеженій ділянці поблизу овального віконця. Відповідно буде збуджуватися і менша кількість рецепторних клітин — лише ті, що розміщені на базилярній мембрані біля основи завитки.
Аналізатор слуху. Від рецепторних клітин спірального (кортієвого) органу подразнення передається клітинам спірального ганглію. Аксони цих клітин входять до складу волокон завиткового нерва, який у внутрішньому слуховому проході з'єднується з вестибулярним нервом в один статоакустичний нерв. Після входу в черепну порожнину нервові волокна, що належать клітинам спірального ганглію, відокремлюються від статоакустичного нерва і входять у довгастий мозок, закінчуючись на нейронах слухового горбика. Клітини слухового горбика є другими нейронами слухового аналізатора і посилають відростки до медіальних колінчастих тіл зорових горбів, де розміщуються мультиполярні клітини (треті нейрони), аксони яких досягають клітин кори великих півкуль головного мозку.