ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 4. СПЕЦІАЛЬНА ГІСТОЛОГІЯ
4.5 Ендокринна система (ендокринні залози)
4.5.2 Периферійні ланки ендокринної системи
Щитоподібна залоза (від лат. glandula thyreoidea).
Перенхіма щитоподібної залози містить клітини двох типів: тироцитів та паратироцитів.
Тироцити розвиваються із вип'ячування ентодермального епітелію вентральної стінки передньої кишки між першою та другою парами зябрових кишень. Епітелій утворює пухкі сіті трабекул, які галузяться. З них формуються замкнені фолікули, а в проміжки між останніми вростає мезенхіма із кровоносними судинами та нервами. Мезенхіма утворює строму.
Ііаратироцити - парафолікулярні клітини, розвиваються з нейробластів нервового гребеня. Вони залягають поодинці в стінці фолікула між базальною основою тироцитів і базальною мембраною, проте не досягають своєю апікальною поверхнею просвіту фолікула. Крім того, парафолікулярні клітини знаходяться в міжфолікулярній сполучній тканині. їх характеристика буде наведена нижче.
Зовні щитоподібна залоза обмежена сполучнотканинною капсулою, її прошарки спрямовані всередину й поділяють орган на частки. Структурно-функціональною одиницею щитоподібної залози є фолікул. Це мікроскопічний міхурець, стінка якого утворена одним шаром клітин — тироцитів, що лежать на базальній мембрані.
Фолікули розділяються прошарками пухкої сполучної тканини, які супроводжуються кровоносними та лімфатичними капілярами, що обплітають фолікули, а також нервові волокна. В сполучнотканинних прошарках знаходяться групи компактних тироїдних епітеліальних клітин.
Форма тироцитів тісно пов'язана з функціональною активністю щитоподібної залози. При помірній функціональній активності тироцити мають кубічну форму та кулясті ядра. Колоїд, що утворюється тироцитами, у вигляді гомогенної маси заповнює просвіт фолікула. На апікальній поверхні тироцитів містяться мікроворсинки. Бічні поверхні сусідніх тироцитів формують десмосомні контакти. Плазмолема базальної поверхні цих клітин утворює інвагінації.
Коли функціональна активність тироцитів підвищується, вони набувають призматичної форми, зростає кількість і висота мікро- ворсинок. Інтрафолікулярний колоїд при цьому стає рідким і містить численні резорбційні вакуолі. Одночасно базальна поверхня тироцитів набуває складчатості, що збільшує стикання тироцитів з перифолікулярним простором. При гіпофункції щитоподібної залози висота тироцитів зменшується, вони стають плоскими, а їх ядра набувають витягнутої форми, діаметр фолікулів збільшується, колоїд стає ущільненим.
Цитоплазма тироцитів містить добре розвинені мембранні структури гранулярної ендоплазматичної сітки, комплекс Гольджі, мітохондрії, тобто органели, що мають відношення до білкового синтезу.
Секреторна функція тироцитів складна й відбувається у дві фази: продукції та виведення гормонів. Фаза продукції полягає у поглинанні через базальну поверхню амінокислот, у тому числі тирозину, йоду та інших мінеральних іонів, деяких вуглеводів, води з крові. У ендоплазматичній сітці синтезуються молекули тироглобуліну. Утворені сполуки поступово переміщуються у зону комплексу Гольджі, де до поліпептидної основи приєднуються вуглеводні компоненти і утворюються везикули, що містять тироглобулін.
Цей процес завершується в апікальній частині тироцита, звідки тироглобулін шляхом екзоцитозу надходить всередину фолікула, нагромаджуючись у вигляді колоїду (рис. 121,122).
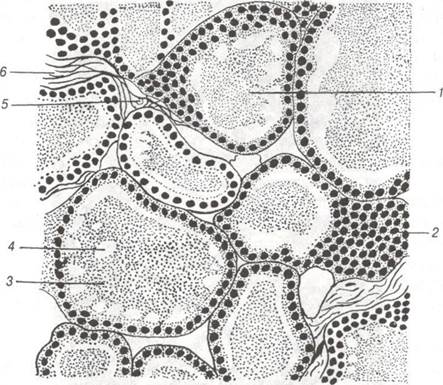
Рис. 121. Щитоподібна залоза коня:
1-фолікул; 2-стінка фолікула; 3-колоїд; 4-вакуоля; 5-капіляр; 6-сполучна тканина.
Йод поглинається тироцитами з крові у вигляді іонів, які під впливом ферменту пероксидази перетворюються у атомарний. Це відбувається на апікальній поверхні тироцита та його мікроворсинок. До тирозинів, що знаходяться у складі тироглобіну, приєднується спочатку по одному атому йоду, утворюються монойодтирозини, потім по другому, і монойодтирозини переходять у дийодтирозини, які об'єднуються у тетрайодтиронін (тироксин). Поряд з останнім утворюється і трийодтиронін.
Фаза виведення. Тироцити своєю апікальною поверхнею шляхом ендоцитозу фагоцитують частини інтрафолікулярного колоїду, які всередині цитоплазми перетворюються у внутрішньоклітинні краплі
колоїду. Лізосоми з'єднуються з ними, після їх розщеплення утворюються тиреоїдні гормони, які через базальну частину тироцита надходять у загальний кровообіг.
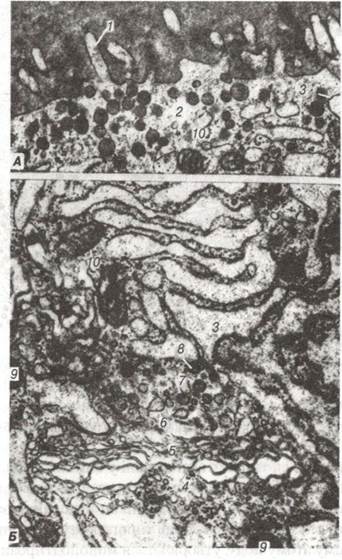
Рис. 122. Електронограма фолікулярної клітини щитоподібної залози:
А- апікальна частина клітини: 1-мікроворсинки; 2-апікальні гранули; Б — органели, які беруть участь у секреції тиреоглобуліну:
3-розтягнуті цистерни гранулярної ендоплазматичної сітки; 4-комплекс Гольджі; 5-транспортні міхурці; 6-пресекреторні гранули;
7-секреторні гранули; 8-облямовані міхурці; 9-лізосоми; 10-мітохондрії.
Паратироцити (кальцитоніноцити - клітини-«К») мають більший розмір ніж тироцити, полігональну або неправильну форму. У цитоплазмі кальцитоніноцитів добре розвинена гранулярна ендоплазматична сітка, комплекс Гольджі. На відміну від тироцитів, вони не поглинають йод. Паратироцити утворюють нейроаміни (норадреналін та серотонін) і синтезують олігопептидні гормони — кальцитонін та соматостатин. Існують два різновиди клітин. У перших переважають дрібні сильно осміофільні гранули. Ці клітини продукують кальцитонін. Цитоплазма другого типу клітин містить великі, проте слабо осміофільні гранули. Ці клітини продукують соматостатин.
Щитоподібна залоза іннервується волокнами симпатичної та парасимпатичної нервової системи. Кровопостачання відбувається за рахунок гемокапілярів, що відходять від сонної артерії.
Прищитовидна залоза — відіграє важливу роль у регулюванні метаболізму кальцію. Вона розвивається як потовщення епітелію третьої та четвертої пар зябрових кишень. Зовні паращитовидна залоза вкрита сполучнотканинною капсулою, остання найбільшого розвитку досягає у великої рогатої худоби та коней. Залежно від співвідношення паренхіми та сполучної тканини, у паращитовидній залозі сільськогосподарських тварин виділяють такі архітектонічні типи.
1. Компактний — орган складається їз суцільної маси паренхіматозних клітин: тонкі сполучнотканинні елементи супроводжують лише кровоносні судини, проте розчленування вони не викликають. Цей тип характерний для собак.
2. Сітчастий тип — сполучнотканинні перекладини та тяжі паренхіми мають сіткоподібне розміщення: зустрічається у коней та дрібних жуйних.
3. Часточковий — залозисті клітини утворюють округлі часточки, які повністю обмежені сполучною тканиною; він характерний для паращитовидних залоз свині.
4. Губчастий тип — характеризується значним розвитком сполучнотканинних прошарків; інколи спостерігається у коней та великої рогатої худоби.
Перенхіма прищитовидної залози складається із трабекул-епітеліальних тяжів або скупчень епітеліальних ендокринних клітин — паратироцитів, відокремлених тонкими прошарками пухкої сполучної тканини з численними капілярами.
Розрізняють головні та оксифільні паратироцити. Головні паратироцити містять базофільну цитоплазму, периферія її багата на полісоми, добре розвинений комплекс Гольджі, мітохондрії.
При посиленні секреторної активності прищитовидної залози головні клітини збільшуються у розмірі. Серед головних паратироцитів розрізняють світлі та темні. Цитоплазма світлих паратироцитів містить глікоген. Вважається, що головні, світлі та оксифільні паратироцити не є самостійними різновидами паратироцитів, а лише віковим або функціональним станом клітин.
Паратироцити продукують білковий гормон — паратирин, який впливає на кісткову тканину — депо нагромадження кальцію в організмі, стимулює звільнення останнього, що призводить до підвищення рівня цього іона в крові, а в кістках настає часткова демінералізація.
Таким чином, паратирин та кальцитонін парафолікулярних клітин щитовидної залози є антагоністами. Прищитовидна залоза за принципом зворотного зв'язку реагує на коливання рівню кальцію в крові. Паратироцити мають рецептори, здатні безпосередньо сприймати прямий вплив іонів кальцію на них. Іннервуються паращитовидні залози симпатичними та парасимпатичними нервами.
Надниркові залози є парними органами. До їх складу входить кіркова (інтерреналова) та мозкова (супрареналова) частини, об'єднані в один орган.
Кіркова частина розвивається з целомічного епітелію. Ці клітини проникають у підлеглу мезенхіму і формують тяжі. їх великі оксифільні клітини утворюють первинну (фетальну) кору. В подальшому первинна кора обростає дрібними базофільними клітинами целомічного епітелію кореня брижі, з яких утворюється дефінітивна кора. Фетальна кора залишається до народження, потім зникає.
Мозкова частина розвивається з клітин нервового гребеня, які мігрують у вентральному напрямку, стають залозистими. Ці клітини споріднені з солями хромової кислоти, тому їх називають хромофінними. Найбільша їх частина проникає у центральну закладку кори і перетворюється у мозкову речовину. У зародка хромофінні клітини спочатку містять лише норадреналін, а адреналін з'являється на пізніших стадіях ембріогенезу.
Зовні надниркові залози обмежені сполучнотканинною капсулою, її зовнішня частина щільна, а внутрішня пухкіша. Від капсули всередину органа проходять тонкі прошарки, які поряд з капілярами та нервами залягають між тяжами паренхіми. Під капсулою знаходиться тонкий прошарок дрібних епітеліальних клітин, за їх рахунок регенерує кіркова речовина.
Кіркова частина утворена поверхневою — дуговою, середньою — пучковою і глибокою — сітчастою зонами.
Дугова зона утворена клітинами, переважно, циліндричної форми з овальними або круглими ядрами. Ці ендокриноцити вклинюються між собою і утворюють дугоподібні структури, що характерно для надниркових залоз коней, в меншій мірі — для інших тварин. Цитоплазма цих клітин містить агранулярну ендоплазматичну сітку у вигляді дрібних міхурців, між якими знаходяться рибосоми, добре розвинений комплекс Гольджі та мітохондрії. Ендокриноцити дугової зони продукують гормон альдостерон — мінералокортикоїдний гормон, що регулює вміст натрію в організмі, крім того, мінералокортикоїди посилюють запальні процеси.
Між дуговою та пучковою зонами знаходиться вузька смужка дрібних малодиференційованих клітин. Її називають проміжною або суданофільною зоною. Ці клітини вважаються джерелом фізіологічної регенерації пучкової та сітчастої зон.
Пучкова зона найбільша. Ендокриноцити, що входять до її складу, мають великі розміри, кубічну або призматичну форму, утворюють тяжі. На поверхні, що межує з капілярами, знаходяться мікроворсинки. Цитоплазма цих клітин багата на краплі ліпідів. Мітохондрії великі, з кристами у вигляді звивистих трубок, які розгалужуються. Добре розвинена агранулярна ендоплазматична сітка, мають місце рибосоми.
Серед ендокриноцитів поряд з світлими мають місце темні, з ущільненою цитоплазмою та незначним вмістом ліпідних включень, проте з підвищеним рівнем рибонуклеопротеїдів. Темні клітини містять розвинену агранулярну та гранулярну ендоплазматичні сітки. Світлі та темні клітини є різновидами функціонального стану ендокриноцитів.
Ендокриноцити пучкової зони синтезують глюкокортикоїдні гормони (кортизон, гідрокортизон, кортикостерон), які регулюють обмін вуглеводів, білків, ліпідів, стимулюють енергетичний обмін, пригнічують запальні процеси.
Ендокриноцити сітчастої зони кубічної, полігональної або округлої форми, утворюють розгалужені пучки. Кількість темних клітин збільшується, а ліпідних речовин зменшується. Клітини сітчастої зони синтезують андрогенстероїдний гормон, подібний до тестостерону, частково — естрогени та прогестерон (рис. 123).
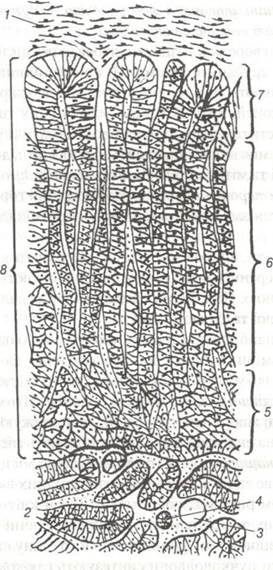
Рис. 123. Надниркова залоза коня:
1-капсула; 2-мозкова речовина; 3-кровоносна судина; 4-нерв; 5-сітчаста зона; 6-пучкова зона; 7-клубочкова зона; 8-кіркова речовина.
Мозкова речовина міститься у центральній частині наднирника. До її складу входять хромофінні клітини, симпатичні нейрони, нервові волокна та синусоїдні капіляри. Хромофінні клітини великого розміру, округлої або полігональної форми. Серед них розрізняють епінефроцити та норепінефроцити. Епінефроцити — світлі клітини, містять секреторні гранули, продукують адреналін. Цитоплазма норепінефроцитів має темний вигляд, містить секреторні гранули гормону норадреналіну. Норадреналін та адреналін відносять до катехоламінів. Адреналін посилює роботу серця, бере участь в регуляції вуглеводного обміну. Норадреналін є медіатором нервового збудження, звужує стінку кровоносних судин та підвищує кров’яний тиск, впливає на нейросекреторну функцію гіпоталамуса.
Незначні скупчення клітин мозкової речовини наднирника можуть знаходитися поряд із симпатичними гангліями, тому їх називають парагангліями.
Васкуляризація мозкової та кіркової речовини наднирника здійснюється загальним кровопостачанням. Артерії, що надійшли до надниркових залоз, розгалужуються на артеріоли, що утворюють густу субкапсулярну сітку; від неї відходять капіляри у кіркову частину. В сітчастій зоні капіляри входять у мозкову речовину, де набувають вигляду синусоїдів і зливаються у венули, які переходять у венозні сплетення мозкової речовини. Поряд з ними у мозкову речовину входять також артерії. У мозковій частині розгалуження кровоносних судин таке, що кожний хромафіноцит одним кінцем контактує з артеріальним капіляром, а другим — з венозним синусоїдом. Кров із кори та мозкової речовини надходить у воротну вену печінки.
Іннервуються надниркові залози нервовими імпульсами по черевних та блукаючих нервах.