ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 4. СПЕЦІАЛЬНА ГІСТОЛОГІЯ
4.5 Ендокринна система (ендокринні залози)
4.5.1 Центральні регуляторні утворення ендокринної системи
Гіпоталамус (hypothalamys) — вищий центр ендокринних функцій. Він контролює і об'єднує усі вісцеральні функції організму та ендокринні механізми регуляції з нервовими. Займає базальну частину проміжного мозку і оточує своєю речовиною нижню половину третього мозкового шлуночка. У сірій речовині гіпоталамуса знаходиться 32 пари ядер, розміщених в передній, середній і задній зонах.
У передній зоні знаходяться дві пари ядер, що складаються із великих пептидохолінергічних нейросекреторних клітин. Це супраоптичні та паравентрикулярні ядра. Їх клітини мають добре розвинені органели — гранулярну ендоплазматичну сітку, комплекс Гольджі, вони забезпечують синтез і виділення гормонів. У цитоплазмі нейросекреторних клітин виявляють специфічні гранули, що містять підготовлені до виведення біологічно активні речовини.
Клітини супраоптичних ядер продукують гормон вазопресин, він викликає скорочення міоцитів судин, чим зумовлює підвищення тиску крові; регулює обмін води, оскільки впливає на зворотне всмоктування води
в сечових канальцях нирок. З урахуванням цієї дії вазопресин називають антидіуретичним гормоном. Також важливу роль відіграє вазопресин у регуляції температури тіла, діяльності серцево-судинної системи.
Нейросекреторні клітини паравентрикулярних ядер синтезують окситоцин, який сприяє скороченню міоцитів матки та молочної залози. Гормони супраоптичних та паравентрикулярних ядер по аксонах нейросекреторних клітин нагромаджуються у задній частці гіпофіза, потім надходять у кров.
Середня зона гіпоталамуса містить дрібні пептидоадренергічні клітини, вони об'єднуються в аркуатне, дорсомедіальне, вентромедіальне, супрахиазматичне ядра та преоптичну зону. Клітини цих ядер синтезують біологічно активні речовини, названі ліберинами і статинами, які впливають на клітини передньої частки гіпофіза. Ліберини і статини об'єднують спільною назвою — релізінг-факторів. Ліберини стимулюють продукцію і виведення у кров гормонів гіпофіза до секреції відповідних тропних гормонів, які впливають на клітини-цілі. Крім того, гіпоталамус надсилає свої імпульси до регульованих ефекторів прямо по симпатичних або парасимпатичних нервах останніх, без опосереднення гіпофізом, тобто парагіпофізарно.
Нейросекреторна діяльність гіпоталамуса, в свою чергу, зазнає впливу вищих відділів головного мозку, особливо лімбічної системи, мигдалевих ядер, гіпокампа та епіфіза.
Гіпофіз (від гр. hypophysis cerebri- під, phyomai — рости) належить до центральних регуляторних органів ендокринної системи. Гіпофіз продукує гормони, дії яких спрямовані на регуляцію функції периферійних залоз ендокринної системи, а також на обмін речовин клітин організму неендокринної природи.
Розвиток гіпофіза відбувається за рахунок взаємодії двох ембріональних зачатків — епітеліального та нейрального. Із ектодермального епітелію, що вистеляє ротову ямку зародка, утворюється гіпофізарна кишеня, вона розвивається у напрямку закладки головного мозку і дає початок аденогіпофізу. Назустріч гіпофізарній кишені з боку проміжного міхура зародка головного мозку формується виріст, який перетворюється у лійку третього шлуночка мозку. Нейроглія дистального кінця лійки внаслідок розростання утворює нейрогіпофіз. Проксимальна частина лійки звужується і стає гіпофізарною ніжкою, вона зв'язує гіпоталамус з гіпофізом (рис. 117).
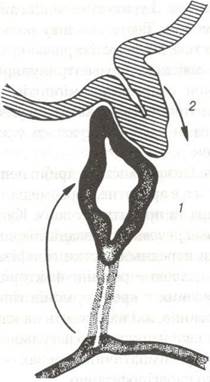
Рис. 117. Схема виникнення зародка гіпофіза із двох його головних джерел:
1-випин даху ротової бухти (гіпофізарна кишеня Ратке); 2-нижній відділ третього мозкового шлуночка.
Диференціювання епітеліальної гіпофізарної кишені починається розростанням передньої стінки гіпофіза, яка перетворюється в передню частку гіпофіза: задня стінка цієї кишені залишається вузькою середньою (проміжною) часткою.
Між передньою та середньою частками (за винятком гіпофіза коня) зберігається залишок порожнини гіпофізарної кишені у вигляді вузької гіпофізарної щілини. Таким чином, ембріональний розвиток зумовив будову гіпофіза. Він складається із двох частин: адено- та нейрогіпофіза.
До складу аденогіпофіза входять передня (дистальна), проміжна та туберальна частки. Четвертою часткою гіпофіза (рис. 118) є нейрогіпофіз, який за походженням є нейрогліальним.
Передня частка утворена розгалуженими епітеліальними тяжами — трабекулами, проміжки між останніми містять пухку сполучну тканину та синусоїдні капіляри. Трабекули утворені залозистими клітинами-аденоцитами, серед яких мають місце три різновиди.
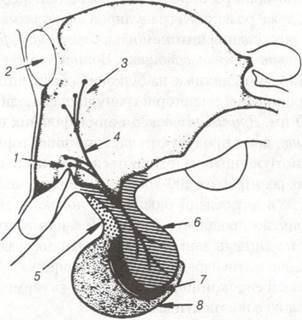
Рис. 118. Схема гіпоталамо-нейрогіпофізарного тракту:
1-супраогітичне ядро; 2-передня комісура; 3-паравентрикулярне ядро; 4-гіпоталамогіпофізарний тракт;
5-горбкова частина; 6-задня частина; 7-проміжна частка; 8-передня частка.
Одні з них знаходяться на периферії трабекул, у цитоплазмі містять секреторні гранули, які інтенсивно сприймають барвники. Ці аденоцити називають хромофільними ендокриноцитами. Вони становлять до 40% клітинної маси передньої частки аденогіпофіза.
Інші клітини займають середню частину трабекули й не мають чітких меж. їх цитоплазма не містить гранул, слабо сприймає барвники, тому такі клітини називають хромофобами. Хромофобних клітин близько 60%.
Хромофільні аденоцити здатні сприймати або кислі барвники, або лужні, тому перші відносять до ацидофільних, другі — до базофільних.
Найбільший розмір мають базофільні аденоцити, середній — ацидофільні, найменші — хромофобні клітини.
Ацидофільні ендокриноцити округлої або овальної форми, розміщуються групами. В їх цитоплазмі знаходяться великі щільні білкові гра
нули, що сприймають кислі барвники — еозин, кислий фуксин, оранж Ж. Центральну частину ацидофільних ендокриноцитів займає ядро, до якого прилягає помірно розвинений комплекс Гольджі, мітохондрії великого розміру, дуже розвинена гранулярна ендоплазматична сітка, що має паралельне розміщення цитомембран. Серед ацидофільних ендокриноцитів розрізняють соматотропоцити. Вони продукують соматотропний гормон (СТГ), який впливає на білковий обмін, чим стимулює ріст тіла. У соматотропоцитах секреторні гранули шароподібної форми, а їх діаметр 300-400 нм. Другим різновидом ацидофільних ендокриноцитів є мамотропоцити, вони продукують лактотропний гормон — ЛТГ, або пролактин. Мамотропоцити відрізняються великими цитоплазматичними гранулами, розмір яких 400-700 нм. Основне значення ЛТГ (про- лактину) полягає в активізації біосинтезу молока в молочній залозі. Крім того, пролактин продовжує функціонування жовтого тіла в яєчнику. За кількістю ацидофільні ендокриноцити досягають 30-36% усіх хромофільних аденоцитів передньої частки гіпофіза.
Усі ацидофільні ендокриноцити продукують гормони, що за своєю хімічною природою є поліпептидами.
Базофільні ендокриноцити гіпофіза містять гранули, що сприймають основні барвники. Відносна кількість цих клітин становить 4-10% від загальної кількості хромофільних аденоцитів передньої частки. Серед базофільних ендокриноцитів розрізняють гонадотропні та тиротропні клітини.
Гонадотропні становлять 2,2-3,5% від загальної кількості клітин аденогіпофіза. Це великі округлі клітини, більшість їх мають електронно негативне зображення добре розвиненого комплексу Гольджі у вигляді кільцеподібної структури, що має назву макула. Секреторні гранули величиною 200-300 нм, сприймають такі барвники, як альціа- новий синій, альдегід-фуксин та ін. Ендоплазматична сітка цих клітин має вигляд невеликих міхурців або вузьких канальців. Мітохондрії паличкоподібні, а їх матрикс має підвищену електронну щільність.
Гонадотропоцити синтезують фолікулостимулюючий гормон (ФСГ-фолітропін), який впливає на проліферацію сперматогоній сім'яників та фолікулярних клітин яєчника. Другий — лютеїнізуючий гормон (ЛГ-лютропін), його функція полягає у стимуляції жовтого тіла яєчника та стимуляції статевих гормонів самця інтерстиціальни- ми клітинами сім’яника (рис. 119).
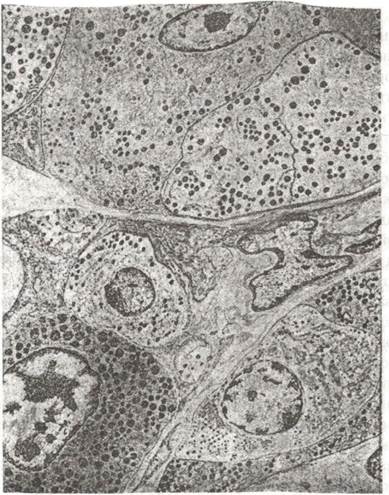
Рис. 119. Електронограма передньої частки гіпофіза х 5450:
а-соматотропоцит; б-мамотропоцит; в-кортикотропоцит.
Тиротропні клітини характеризуються кутоподібною або неправильною формою. Цитоплазма тиротропоцитів містить чітку грануляцію діаметром 80-150 нм, яка сприймає альдегід-фуксин, альдегід-тіонин, альціановий синій та інші барвники. Кількість тиротропоцитів порівняно невелика й становить у пацюків приблизно 1,8-2,9% від загальної кількості секреторних клітин аденогіпофіза. Тиротропоцити продукують тиротропний гормон — тиротропін, який впливає на щитоподібну залозу.
Крім названих ацидофільних та базофільних ендокриноцитів, передня частка гіпофіза містить третю групу хромофільних клітин, названих кортикотропоцитами. Ці клітини займають переважно центральну зону передньої частки гіпофіза. Вони продукують білковий адренокортикотропний гормон — АКТГ-кортикотропін, який підвищує гормоноутворюючу функцію кіркової частини наднирників. Кортикотропоцити мають неправильну багатокутну форму, їх ядра складаються із окремих часток. Мітохондрії та ендоплазматична сітка добре розвинені. Секреторні гранули мають розмір 100-200 нм і вигляд мембранних міхурців з щільною серцевиною. Між мембраною і серцевиною залишається світлий простір.
Вважається, що група хромофобних ендокриноцитів є збірною і містить аденоцити з різним ступенем розвитку, диференціювання та різного фізіологічного значення.
Серед хромофобних клітин знаходяться, так звані, фолікулярно-зірчасті клітини, проте їх природа та значення не з'ясовані. Ці клітини здатні формувати мікрофолікулярні структури з відкладенням у них секрету.
Проміжна частка гіпофіза в препаратах, фарбованих гематоксиліном та еозином, має вигляд базофільної смужки епітелію. Її клітини здатні продукувати білковий або слизовий секрет, який нагромаджується, утворюючи фолікулярні кісти. Функціональне значення проміжної частки гіпофіза достовірно установлено лише для нижчих пойкілотермних хребетних тварин, у яких меланостимулюючий гормон (МСГ) має провідне значення у регуляції забарвлення тіла. Проміжна частка аденогіпофіза містить меланоцитостимулюючий гормон (меланоцитотропін), а також ліпотропін — гормон, що бере участь у метаболізмі ліпідів. Є припущення про те, що меланоцитотропін, ліпотропін, а також АКТГ утворюються у головному мозку шляхом розщеплення великої початкової молекули церебрального пептиду на менші фрагменти, кожний із яких стає одним із названих гормонів.
Туберальна частка аденогіпофіза — відділ, що межує з гіпофізарною ніжкою та медіальним підвищенням гіпоталамуса. Вона утворена епітеліальними тяжами кубічних клітин із помірно базофільною цитоплазмою. Від останніх, як і від епітелію проміжної частки, у передню частку відходять трабекули; у деяких клітинах туберальних тяжів виявляють базофільні гранули. Функція клітин туберальної частки гіпофіза не визначена.
Гіпоталамо-аденогіпофізарне кровопостачання здійснюється портальною (ворітною) судинною системою. Приносні гіпофізарні артерії вступають в медіальне підвищення медіального гіпоталамуса і утворюють сітку капілярів (первинне капілярне сплетення портальної системи), з якою контактують кінцеві відділи аксонів нейросекреторних клітин адеиогіпофізарної зони гіпоталамуса.
Капіляри первинного сплетення зливаються у портальні вени, які розміщуються вздовж гіпофізарної ніжки до аденогіпофіза, де розпадаються на капіляри синусоїдного типу (вторинна капілярна сітка) і розгалужуються між трабекулами паренхіми залози.
Синусоїди вторинної капілярної сітки зливаються у виносні вени, в яких кров, збагачена на гормони передньої частки, надходить у загальну циркуляцію.
Нейрогіпофіз {задня частка) утворена клітинами епендими, які називають пітуїцитами. За формою вони веретеноподібні або зірчасті, мають численні тонкі відростки, що досягають адвентиції кровоносних судин або базальної мебрани капілярів.
Нейрогіпофіз акумулює вазопресин та окситоцин, які продукуються великими пептидохолінергічними нейросекреторними клітинами переднього гіпоталамуса.
Аксони цих нейросекреторних клітин досягають нейрогіпофіза, закінчуються великими терміналями, названі тільцями Херінга, де нагромаджуються секреторні гранули окситоцину та вазопресину. Терміналі контактують з капілярами, у які й надходять гормони.
Епіфіз (від гр. ері - над, phyomai — рости) (шишкоподібне тіло) один із центральних органів ендокринної системи.
Знаходиться епіфіз між півкулями великого мозку та мозочком. Зовні він обмежений сполучнотканинною капсулою, її прошарки проникають всередину органа, розгалужуються і поділяють його паренхіму на часточки. Паренхіма епіфіза містить секретоутворюючі клітини — пінеалоцити та гліальні клітини — гліоцити, які відносять до астроцитної глії.
Гліоцити виконують опорну функцію, їх відростки мають тісний зв'язок з сполучнотканинною стромою.
Пінеалоцити займають центральну частину часток, вони більші від гліоцитів, мають полігональну форму, світле ядро з великими ядерцями. Від пінеалоцитів відходять розгалужені довгі відростки, які тісно контактують з відростками гліоцитів. Відростки пінеалоцитів біля капілярів утворюють булавоподібні розширення, у складі яких містяться осміофільні гранули. Серед пінеалоцитів виділяються світлі пінеалоцити з бідною на секреторні включення гомогенною цитоплазмою; та темні пінеалоцити меншого розміру з ацидофільними або базофільними гранулами в цитоплазмі. Вважають, що світлі та темні пінеалоцити не самостійні різновиди, а являють собою клітини, що знаходяться у різному функціональному стані. Цитоплазма пінеалоцитів містить численні мітохондрії, розвинений комплекс Гольджі, лізосоми, міхурці агранулярної ендоплазматичної сітки, рибосоми та полісоми. Гліальні клітини, переважно, знаходяться на периферії часток. їх цитоплазма без видимих структур, ядра ущільнені. Під електронним мікроскопом у цитоплазмі виявляють мікрофіламенти діаметром 5-6 нм.
Поряд з пінеалоцитами і гліоцитами зустрічаються фіброцити, макрофаги, лаброцити, лімфоцити, плазмоцити та пігментні клітини.
Функція епіфіза складна й різноманітна. Він затримує розвиток статевої системи, гальмує статеву функцію. Остання зумовлюється тим, що пінеалоцити продукують серотонін, який в епіфізі перетворюється у мелатонін. Як вважають, цей нейроамін пригнічує секрецію гонадоліберину гіпоталамусом та гонадотропінів передньої частки гіпофізів. В той же час пінеалоцити продукують ряд білкових гормонів, зокрема антигонадотропін, який послаблює секрецію лютропіну передньої частки гіпофіза. Поряд з антигонадотропіном пінеалоцитами синтезується значна кількість регуляторних пептидів, яка досягає 40.
Епіфіз забезпечує регуляцію фотоперіодичності роботи органів і систем організму, його циркадних ритмів — коливання активності клітин у зв'язку із зміною дня та ночі. Механізм реагування епіфіза на зміни освітленості пов'язаний із сприйняттям ним подразнень від сітківки ока по симпатичних нервових стовбурах.
Як показують порівняльно-анатомічні дані, епіфіз виникає філогенетично з тім 'яного ока, яке до цього часу існує в функціональному стані у новозеландської ящірки - гаттерії. Воно затягнене тонкою шкірою, містить сітківку, рогівку та кришталик.(рис. 120).
Пінеальна залоза виникає у вигляді вип’ячування та виросту дорсальної стінки третього мозкового шлуночка. Зачаток епіфіза у великої рогатої худоби з'являється вперше у 7-тижневих зародків, а паренхіма відмежовується після 21-тижневого ембріонального віку.
Вікова інволюція епіфіза настає у ранньому віці. Частина пінеалоцитів при цьому атрофується, строма розростається, у ній нагромаджуються вапняні нашарування — мозковий пісок.

Рис. 120. Епіфіз:
1-пінеалоцит; 2-булавоподібні розширення відростків пінеалоцитів.