Основи еволюції - Корж О.П. - 2006
Частина I. МІКРОЕВОЛЮЦІЯ
Розділ 6. ІЗОЛЯЦІЯ
6.2. Типи ізоляції
Явище ізоляції надзвичайно різноманітне, тому не завжди вдається встановити справжні причини походження тієї чи іншої її форми. Обмеження схрещуваності особин може бути повним унаслідок дії будь-якої різкої ізоляції або частковим, коли розмежованість між новою та вихідною формами встановлюється лише поступово. Крім обмежень схрещуваності в просторі, можливе обмеження ще й у часі. Окремі вчені говорять про первинні та вторинні форми ізоляції, що додатково ускладнює класифікацію цих явищ.
Географічна ізоляція як розмежування особин у просторі. Під географічною ізоляцією розуміють перебування особин у різних ареалах, через що вони не можуть зустрітися. З її дією не пов'язана наявність у вихідних форм обов'язкових біологічних відмінностей — вони можуть з'явитися пізніше.
Цей тип ізоляції властивий майже всім організмам, бо існують передумови для його формування: часткове обмеження схрещування досягається вже характером розміщення окремих особин у просторі, за винятком хіба що вузько ендемічних форм, які мають зовсім малий ареал, через що панміксія може бути майже повного. Останні й виникають передусім унаслідок тривалої дії географічної ізоляції. Науці відомо чимало прикладів (Дарвінові в'юрки, гавайські квіткарки, Галапагосові черепахи тощо), коли на островах в умовах ненасичених біо- геоценозів деякі види розвивались шляхом адаптивної радіації (розвиток значної кількості споріднених форм, пристосованих до різних умов існування) з утворенням великої кількості ендемічних видів.
Різні популяції в межах видового ареалу вже з самого початку мають певну географічну ізоляцію, через що обмін генетичним матеріалом між ними ускладнюється. Найбільш яскравим прикладом цього явища слід вважати відносно малорухливі види з
великими ареалами. До них можна віднести звичайну білку (Sciurus vulgaris) або звичайного фазана (Phasianus colchicus), у яких виникає значна кількість підвидів.
Ізоляцію простором пов'язують з репродуктивною активністю тих чи інших організмів, яка зумовлюється рухливістю останніх. Якщо радіус індивідуальної активності малий відносно розмірів популяції й відстані між сусідніми популяціями, то в цьому випадку тиск просторової ізоляції досить відчутний. І, навпаки, значна індивідуальна активність сприяє зменшенню тиску цього фактора. Так, перелітні птахи відзначаються слабкою внутрішньовидовою диференціацією (зокрема, денні хижаки та гусеподібні). Гніздовий консерватизм (лелеки та деякі інші види птахів) також може сприяти формуванню просторової ізоляції.
Більш очевидною є ізоляція бар'єрами, яка полягає у створенні між популяціями певних перешкод географічного (механічного) походження. Розмір необхідних перешкод для задіяння цього елементарного еволюційного фактору багато в чому також залежить від активності тих чи інших видів.
Найбільш типовими прикладами зазначеного є ізоляція водою та гірськими системами для суходільних нелітаючих форм, а для водних - суходолом, морською водою - для прісноводних форм. Але для форм з низькою рухливою активністю навіть невеликі бар'єри можуть стати нездоланними перешкодами.
Виділення в географічній ізоляції наведених вище форм слід вважати умовним, оскільки дія однієї з них часто не лише не виключає, а навіть передбачає дію іншої. Для перелітних птахів відстані, що долаються ними щороку, можна вважати і певним бар'єром, який спричиняє підвищену загибель особин. Для дрібніших форм подібний зв'язок відстані і перешкод стає ще більш очевидним. Так, на острові вулканічного походження Оаху є 25 долин, порослих тропічним лісом і розмежованих між собою скельними хребтами без рослинності. У кожній з них живе свій вид равликів р. Achatinella, що пояснюється неспроможністю останніх перейти з однієї долини до іншої через хребти, які добре прогріваються. Нездатність перебороти цю незначну відстань стала нездоланним бар'єром, унаслідок чого географічна ізоляція зумовила формування 25 незалежних видів.
У житті рослин географічна ізоляція є ще більш складним явищем, оскільки їх рухлива активність обмежується переважно розселенням у фазі насіння чи спор, а розмноження залежить ще й від перенесення пилку запилювачами або вітром. Тому географічна ізоляція рослин значною мірою визначається рухливою активністю тварин-запилювачів.
Особливості біологічної ізоляції
На відміну від географічної ізоляції, пов'язаної з мешканням особин у межах різних ареалів, біологічна полягає в унеможливленні самого процесу схрещування. Усі форми біологічної ізоляції об'єднують у дві великі групи — докогіуляційну та післякопуляційну.
Докопуляційна біологічна ізоляція здійснюється без участі гамет і спрямована на запобігання в той чи інший спосіб процесу копуляції. Механізмів подібної ізоляції розрізняють багато, зокрема ізоляцію в часі, яку можна поділити на фенологічну та чергування поколінь.
Фенологічна ізоляція пов'язана з існуванням так званих ярових та озимих рас, що добре простежується в деяких видів риб. Озимі раси йдуть на місця нересту восени, а ранньою весною в них починається розмноження. Ярові ж раси з настанням весни лише розпочинають переміщення до місць нересту, через що їх процес розмноження в них відбувається значно пізніше. Унаслідок цього вони виявляються ізольованими від попередньої раси в часі. Подібні причини викликають також фенологічні зміни строків у розмноженні птахів та деяких інших організмів.
Однією з форм чергування поколінь є зміна статевого розмноження на вегетативне, що властиве як найпростішим, так багатоклітинним організмам (статевий процес чергується з нестатевим поділом або пупкуванням). Подібна стратегія дозволяє випробувати нові генетичні комбінації, утворені під час статевого процесу, тривалою дією природного добору при вегетативному розмноженні, тобто при стійкому збільшенні кількості особин. Це значно послаблює вплив шкідливих факторів на окремі комбінації й не заважає подальшій мінливості шляхом статевого розмноження.
У багатоклітинних подібні чергування набувають особливого значення, зокрема в разі сидячого способу життя вегетативного покоління (гідроїди, коралові поліпи, мохуватки, деякі асцидії тощо). Інколи подібний тип розмноження встановлюється надовго, що сприяє формуванню великих колоній і запобігає випадковій загибелі певного генотипу. Ще більшого біологічного значення вегетативне розмноження набуває в рослин.
Еволюційні наслідки чергування поколінь полягають у полегшенні фіксування тієї чи іншої генетичної комбінації на перших, найбільш загрозливих етапах завоювання нею права на самостійне існування. Розмноження ж нової форми відбувається після випробування її життєздатності.
Ще більш вагомими в еволюційному значенні є життєві цикли з чергуванням статевих поколінь та нестатевого розмноження (деякі тварини, у першу чергу споровики (рис. 6.1), а також більшість рослин). Ця форма ізоляції не обмежується лише збільшенням кількості копій нової форми, а включає й розвиток гаплоїдних організмів, у яких кожна мутація набуває максимального прояву, через що природний добір має більш жорстку дію. Виходить, що гаплоїдне покоління відіграє роль «санітара», тобто захищає вид від засмічування шкідливими спадковими змінами. Але в ході еволюційного розвитку відбувається зосередження дії природного добору переважно на диплоїдному організмі, в якому краще реалізуються різні ступені прояву та комбінування нових ознак. Особливо наочно це простежується у вищих рослин, у яких найбільшого розквіту набули покритонасінні (спостерігається максимальна редукція гаметофіту).
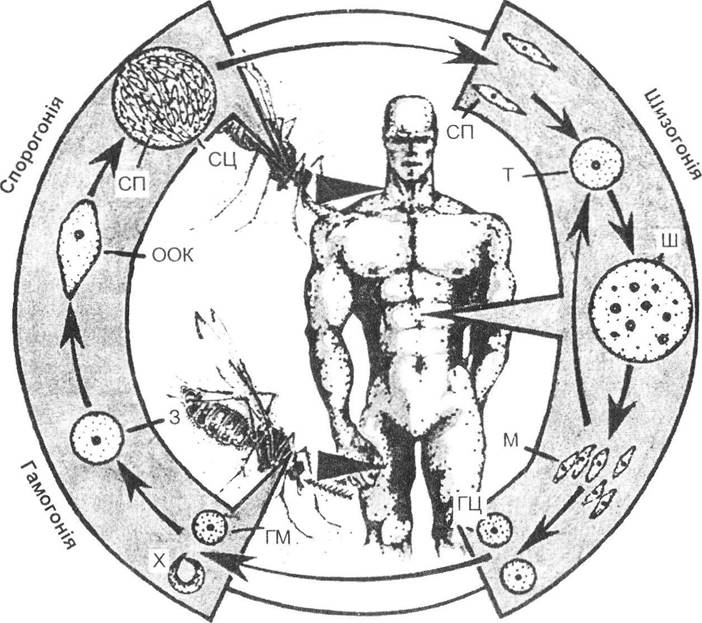
Рис. 6.1. Узагальнена схема циклу розвитку паразитів родини Plasmodium (рис. О.В. Карнаухова):
СП - спорозоїт; Т - трофозоїт; Ш - шизонт; М - мерозоїт; ГМ - гамети; X - копуляція; 3 - зигота; ООК - оокінета; СЦ — спороциста
Близькими за наслідками до попередніх форм ізоляції різних поколінь є здатність до самозапліднення в рослин і партеногенез у тварин. Більшості рослин (за окремими винятками), крім перехресного запліднення, властиве й самозапліднення. Якщо в деяких випадках починає переважати останнє, то це сприяє швидкому поширенню нової форми за умови її життєздатності та конкурентоспроможності. З одного боку, самозапліднення захищає певну генну комбінацію від дезінтеграції, але з іншого організми втрачають пластичність і можуть позбутися здатності до подальшого прогресивного еволюціонування.
Партеногенез у тварин відрізняється тим, що більшість їх форм набуває вигляд гетерогонії (чергування статевого розмноження з партеногенетичним). У коловерток, нижчих ракоподібних (гіллястовусих) та деяких комах (попелиці тощо) розвинулася навіть сезонна цикліка в розмноженні, при якій статеве розмноження відбувається переважно восени з утворенням так званих зимувальних яєць, а впродовж весняно-літнього періоду розвивається кілька партеногенетичних поколінь. У паразитичних форм подібна гетерогонія спричиняє формування складних життєвих циклів, які найбільш добре простежуються на прикладі трематод (рис. 6.2).
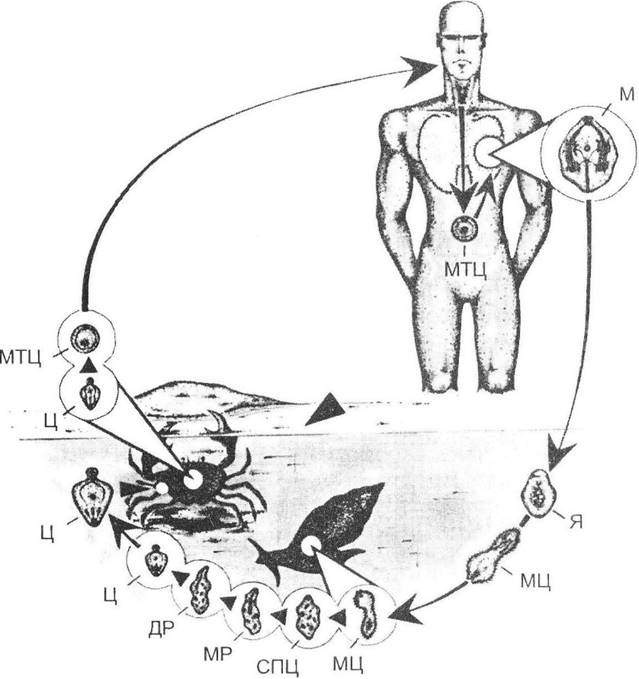
Рис. 6.2. Типовий цикл розвитку трематод на прикладі Paragnimus westermani (рис. О.В. Карнаухова)
М - марита; Я - яйце; МЦ - мірацидій; СПЦ - спороциста; МР - материнська редія; ДР - дочірня редія; Ц - церкарій; МТЦ - метацеркарій
Будь-яка комбінація, що виникає під час схрещування, упродовж літнього сезону здатна розмножитися до розмірів цілої популяції. Як і в усіх попередніх випадках, подібна ізоляція усуває ймовірність випадкової втрати нової форми, хоча перехід до постійного партеногенезу також може завести подальший розвиток групи у безвихідь.
Зменшує ймовірність зустрічі організмів також екологічна ізоляція, яка зумовлюється розходженням кількох близьких груп по різних екологічних нішах, тобто відмінностями у виборі місць для розмноження, використанням певних кормів. При екологічній ізоляції зменшується саме ймовірність зустрічі статевих партнерів, але якщо остання відбувається, то процес репродукції проходить без перешкод.
Спочатку можуть виникати мутації, які створюють генотипи з відмінностями в екології розмноження чи репродуктивній території. Так, у птахів часто з'являються форми, що відрізняються місцями локалізації гнізда в різних частинах гніздової стації (нижче чи вище в кроні дерева тощо).
Багатьом видам властива екологічна ізоляція, пов'язана з тим, що організми надають перевагу різним біотопам. Зокрема, дрохва (Otis tarda) західноєвропейської популяції давно перейшла до гніздування в агроландшафтах. У Казахстані після розорювання цілини та в Росії процеси синантропізації почалися порівняно недавно. На певний період утворилися дві популяції з екологічною ізоляцією між собою: особини першої надають перевагу гніздуванню в умовах цілинних степових ділянок, а другої - в агроландшафтах. Зрозуміло, що в останньому випадку загибель потомства значно більша, ніж у попередньому, але уціліла популяція поступово заміщує вихідну через відсутність або нестачу звичайних місць для розмноження. Екологічні відмінності в біотопах мешкання властиві також козулі (Capreolus capreolus), що освоює агроландшафти, чорному дрозду (Turdus merula), якого можна бачити в міських парках на противагу вихідним біотопам мешкання.
Якщо зустріч організмів можлива в часі й просторі, то включається ще ціла низка ізолюючих механізмів. Серед них етоло- гічна ізоляція, пов'язана в першу чергу з відмінностями в шлюбній поведінці представників різних видів. Вона досить типова для тварин, що пояснюється видоепецифічною шлюбною поведінкою, зрозумілою представникам лише певного виду. Більше того, складні шлюбні ритуали, що складаються з багатьох компонентів, призначені також і для попередження міжвидового схрещування.
Вважається, що саме етологічна ізоляція є основним механізмом, який запобігає утворенню значної кількості міжвидових гібридів тварин, тому вона й має значне поширення.
У кожної тварини існує вроджена здатність як подавати особливі сигнали, властиві лише її виду, так і реагувати виключно на них.
У природі можна спостерігати численні випадки статевої реакції представників одного виду на інших, але дуже рідко вони закінчуються утворенням пар та заплідненням. Так, у метеликів сатира Семела (Eumenis Semele) самці починають залицяння з переслідування статевого партнера в повітрі. Ним може бути метелик іншого виду, якийсь жук, муха чи дрібний птах, аж до листя, що падає з дерев, і навіть власна тінь. Проте лише самиця сатира, якщо вона готова до спаровування, відповідно реагує на переслідування самця - приземляється і не рухається. Інші види переважно тікають. Навіть у випадку схожої поведінки самиці споріднених видів не здатні відтворити всі компоненти шлюбної поведінки, через що спаровування не відбувається.
Подібна картина властива й іншим видам, зокрема плодовим мухам (Drosophila), спаровування між якими унеможливлюється на різних стадіях залицяння через помилки як самця, так і самиці. Шлюбна поведінка риб триголкової (Gasterosteus aculeatus) та дев'ятиголкової (Pungitius pungitius) колючки схожа, дещо відмінним є шлюбний наряд самців та їх поведінка в гнізді під час відкладання ікри, чого вистачає для попередження схрещування (рис. 6.3).
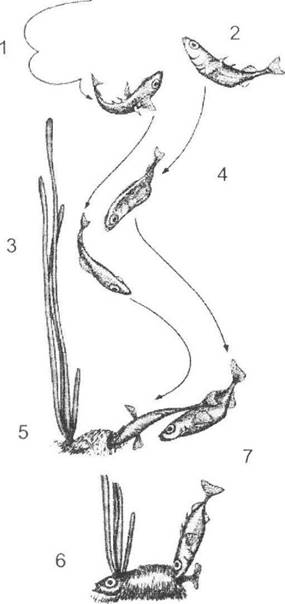
Рис. 6.3. Послідовність залицяння в триголкової колючки (Gasterosteus aculeatus), яка набуває вигляду реакцій за принципом «сигнал - відповідь» (за Н. Тінбергеном, 1993):
1 - зигзагоподібний танець самця; 2 - залицяння самиці; 3 - самець веде самку до гнізда; 4 - вона слідує за ним; 5 - самець показує вхід до гнізда; 6 - самиця входить до гнізда; 7 - самець поштовхує самицю, після чого вона відкладає ікру і випливає
Одним із механізмів формування етологічної ізоляції є імпринтинг (фіксація в пам'яті прикметних ознак об'єктів на ранніх етапах життя) особин свого виду. Зокрема, у Дарвінових в'юрків репродуктивна
ізоляція забезпечується Не тільки відмінностями екологічних ніш, але й специфічною реакцією кожного партнера лише на характерну для його виду форму дзьоба, що пов'язане з особливостями живлення. У деяких випадках, навпаки, імпринтинг може сприяти появі гібридів: при спільному вирощуванні канадської казарки (Branta canadensis) та сірої гуски (Anser anser) можуть утворюватися змішані статеві пари, оскільки вони з дитинства звикають одне до одного.
Особливого значення при формуванні етологічної ізоляції набувають так звані рєлізери - сигнали, що викликають інтенсивну відповідну реакцію в особин свого виду (рис. 6.4). Так, у самців триголкової колючки таким релізером є червона пляма на череві, а в дев'ятиголкової - смолянисто-чорне забарвлення. У багатьох птахів релізерами також виступають яскраві плями в забарвленні пір'яного покриву, розташування, форма і колір яких завжди є видоспецифічними.
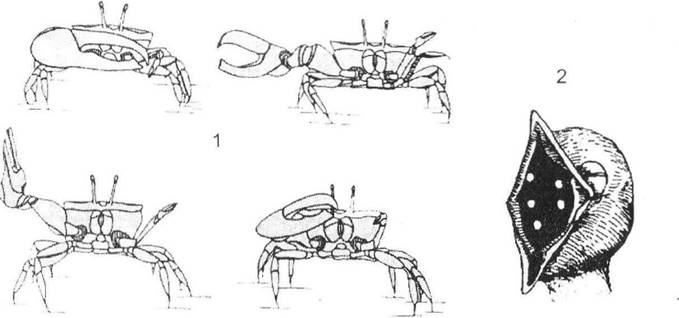
Рис. 6.4. Приклади релізерів, які мають видову специфіку й попереджують міжвидову гібридизацію (за Р. Шовеном, 1972):
1 - самець краба (Uca lactea) використовує клешню як релізер, який свідчить про його стать; 2 - пташеня синиці (Parus major) демонструє батькам кольорові плями в дзьобі (релізери), що стимулює останніх до його годування
Цікавим прикладом етологічної ізоляції є утворення біологічних рас зозулі звичайної (Cuculus canorus), що відрізняються одна від одної генетично закріпленим забарвленням яєць, які схожі на яйця різних видів дрібних горобиних. Репродуктивна ізоляція між ними підтримується видами-хазяями шляхом знищення яєць, які не мають відповідного маскуючого забарвлення. Водночас цей приклад може розглядатись і як екологічна форма ізоляції, оскільки особини займають різні екологічні ніші.
Утруднення схрещування між організмами виникає й внаслідок морфофізіологічної ізоляції. Вона зумовлена окремими мутаціями або морфозами, що виключають можливість перехресного запилення або схрещування. Спочатку особини, що втратили здатність до схрещування, не мали суттєвих генетичних відмінностей. У рослин морфофізіологічна ізоляція залежить від дрібних мутацій, що змінюють швидкість проростання пилку, викликають часткову гетеростилію або інші зміни в будові квітки, що впливає на її взаємовідносини з запилювачами.
У тварин морфофізіологічна ізоляція пов'язана перш за все зі змінами будови чоловічих копулятивних апаратів. Так, у багатьох комах і ссавців будова статевих органів має певні відмінності, що використовується навіть у систематиці (рис. 6.5). Основним змістом подібної ізоляції є неспроможність організмів здійснити статевий акт через невідповідність будови їх статевих органів або ж через відмінності в загальній будові організму. Така ізоляція добре простежується на прикладі дії штучного
добору, унаслідок якої різні породи свійських тварин (зокрема, собаки) не можуть між собою схрещуватися через суттєві відмінності в їх конституції. Таким чином, при морфофізіологічній ізоляції зменшується не ймовірність зустрічі особин одна з одною, а стає неможливим їх запліднення через суттєві морфолого-фізіологічні відмінності.
До ізоляційних механізмів після копуляції належить власне генетична ізоляція, яка може полягати в загибелі зигот після запліднення, розвитку часткової або повної стерильності гібридів чи пониженій їх життєздатності. Найбільш очевидною причиною генетичної ізоляції є суттєві каріотипові відмінності організмів. У рослин цьому сприяє поліплоїдія, у живих організмів у цілому - хромосомні мутації.
Генетична ізоляція може початися загибеллю статевих продуктів після копуляції ще до запліднення. У цьому разі виявляється біохімічна несумісність сперматозоїда з тканинами чужого організму та його ядра з плазмою яйцеклітини. Якщо запліднення все-таки сталося, зародок може загинути на ранніх або пізніших етапах ембріогенезу внаслідок порушення процесів обміну, що лежать в основі онтогенезу та життєвих процесів.
Прикладом останнього типу ізоляції можна вважати американську та європейську норок (Mustela oison і М. lutreola відповідно), гібриди між якими не утворюються через резорбцію ембріонів. Загальновідомими є також приклади гібридів (як штучних, так і природних), які відрізняються стерильністю або пониженою життєздатністю.
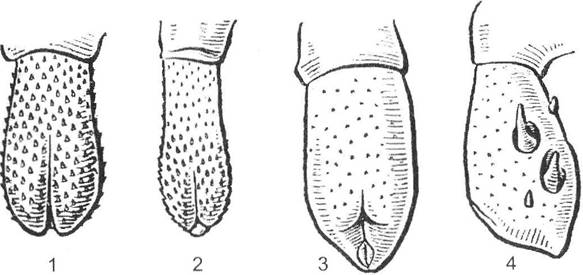
Рис. 6.5. Головки статевих членів самців деяких видів мишівок (за Б.О. Кузнецовим, 1975):
1 - одноколірної (Sicista concolor); 2 - кавказької (S. caucasica); 3, 4 - алтайської рудої (S. napaea) спереду та збоку
За механізмом дії всі форми ізоляції принципово схожі і полягають у закріпленні групових відмінностей унаслідок порушення панміксії у випадку тривалої дії. Первинно можуть виникати будь-які форми ізоляції під впливом зовнішніх стосовно організмів причин (географічна ізоляція) чи внутрішніх (біологічна ізоляція). Наслідком подібного явища завжди є поглиблення відмінностей в утворених популяціях, що може закінчитися врешті-решт генетичною ізоляцією.
Думки вголос
Сучасні наукові дані свідчать на користь того, іцо ізоляція є не лише необхідною, але й обов'язковою передумовою формування нових форм живих організмів. Стануть ці форми новими видами чи навіть набудуть інших систематичних категорій, залежить від багатьох факторів. Ізоляція ж забезпечує саму можливість формування подібної групи. Проте генетична ізоляція стає необов'язковою за умови функціонування інших форм.
Особливого статусу набувають агамні та партеногенетичні форми, кожний представник яких відразу є генетично ізольованим від усіх інших. Таким чином, для цих форм індивідуальні пристосування зрештою можуть набути еволюційного значення.