Основи еволюції - Корж О.П. - 2006
Частина ІІ. МАКРОЕВОЛЮЦІЯ
Розділ 13. Особливості здійснення філогенезу
13.3. Правила еволюції груп
За правилом незворотності Л. Долло (1893), еволюція є необоротнім процесом, і організм не може повернутися до попереднього стану, властивого його предкам. Так, вторинноводні хребетні не перетворюються на риб, а набувають нових пристосувань до водного середовища на своєму рівні організації. При цьому відбувається відновлення структур, які певною мірою лише імітують попередній, більш примітивний стан (плавці, хвіст китоподібних або іхтіозаврів мають зовсім іншу будову порівняно з рибами).
Деякі вчені не погоджуються з наведеним правилом, висловлюючись на користь циклічності еволюційного процесу і можливості повернення до попереднього стану. Справді, зворотні мутації можуть повернути предкову ознаку, але навіть на мік- роеволюційному рівні реконструювати повністю весь генотип неможливо (однаковий генотип мають лише клони). У той же час повернення до так званого дикого типу при формуванні попередніх умов існування свідчить про можливість зворотного розвитку на мікроеволюційному рівні. Визнання оберненості окремих ознак все ж таки не заперечує незворотності еволюційного процесу в цілому.
Правило прогресуючої спеціалізації Ш. Депере (1876) говорить: група, яка стала на шлях спеціалізації, розвивається в подальшому лише з поглибленням цієї спеціалізації. Водночас, на думку автора цього правила, спеціалізація переважно стосується окремих органів (що пов'язане з пристосуванням до специфічних умов існування) і не зачіпає організм у цілому. Так, у птахів, спеціалізованих до польоту, подальша еволюція пов'язана з формуванням різних способів польоту (активний, ширяючий) завдяки поглибленню спеціалізації в будові крила в різних видів.
Інколи до цього ж правила додають закон філогенетичного росту, згідно з яким у кожній філогенетичній гілці простежується тенденція до збільшення розмірів тіла організмів (розвиток спрямований від дрібних представників до більших). Після досягнення критичних розмірів та порушення найважливіших пропорцій відбувається вимирання цієї групи.
За правилом походження від неспеціалізованого предка Е. Копа (1896) нові великі групи походять від порівняно неспеціалізованих представників предкових груп. Саме вони мають вищу потентність до принципово нових пристосувань, що дозволяє їм стати на шлях арогенезу. Так, ссавці походять від найменш спеціалізованих представників рептилій, максимально наближених до котилозаврів. Будь-які спеціалізовані форми не мають необхідних потенцій для подальшого формування нових еволюційних напрямків через наявність великої кількості обмежень.
З правилами Е. Копа та Ш. Депере пов'язана проблема неотенії, про що вже згадувалось раніше. У багатьох спеціалізованих організмів (особливо це стосується сидячих та паразитичних форм) личинки залишаються малоспеціалізованими і, слугуючи для розселення, є активними формами. Тому вважається, що через перехід до неотенії організми здатні знімати обмеження, викликані вузькою спеціалізацією, і йти далі шляхом прогресивного розвитку. Не сперечаючись стосовно поширеності подібних перетворень у природі, зазначимо, що вони не заперечують реальності дії наведених правил.
Правило адаптивної радіації Г.Ф. Осборна (1902): філогенез будь-якої групи супроводжується її розгалуженням на окремі гілки, які розходяться в різних адаптивних напрямках (різні екологічні ніші) від попереднього стану. Це правило повністю узгоджується з принципом дивергенції Ч. Дарвіна і має надзвичайну кількість прикладів (рис. 13.10). Підґрунтям для адаптивної радіації груп є зменшення конкуренції при формуванні кількох видів, які займають різні екологічні ніші (слід ще раз нагадати, що міжвидова боротьба за існування є менш гострою, ніж внутрішньовидова).
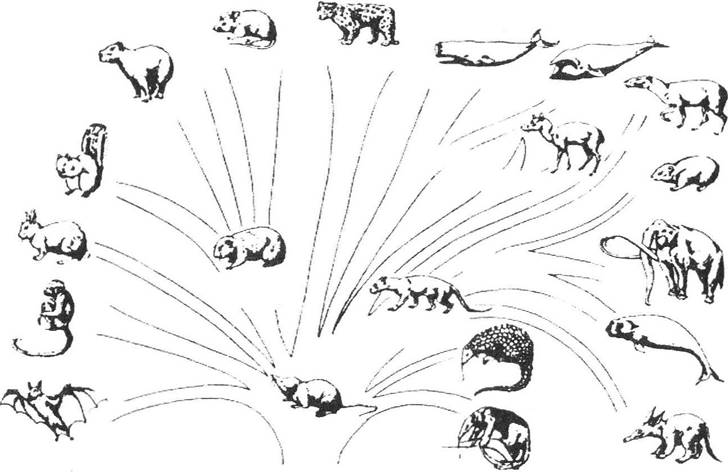
Рис. 13.10. Адаптивна радіація на прикладі плацентарних ссавців (за А. Ромером, Т. Парсонсом, 1992)
Правило чергування головних напрямків еволюції І.І. Шмальгаузена (1939): арогенна еволюція чергується з періодами алогенної еволюції в усіх групах. Необхідність подібного чергування пояснюється тим, що вихід до нової адаптивної зони (арогенез) є досить складним процесом, який супроводжується вимиранням майже всіх проміжних форм. Тому він відбувається лише в разі, коли можливості засвоєної адаптивної зони вже вичерпані, а подальший алогенез стає утрудненим. Вихід же до нової (поки що вільної) адаптивної зони завжди супроводжується видоутворенням з подальшим зайняттям всієї зони.
Правило посилення інтеграції біологічних систем І.І. Шмальгаузена (1961): біологічні системи внаслідок еволюції стають більш інтегрованими, відбувається ускладнення механізмів, які забезпечують цю інтеграцію (гомеостаз екосистем та механізми його підтримання). Це правило свідчить про неможливість зведення біологічних систем будь-якого рівня організації до простої сукупності їх складових. Весь хід еволюційного процесу підтверджує це правило як на онтогенетичному рівні (автономізація та каналізація онтогенезу), так і на популяційно-видовому (складна структура виду) та екосистемному рівні (взаємозв'язок та взаємозалежність усіх представників відповідних екосистем).
Перелік еволюційних правил можна було б продовжити, але й наведені мають певні винятки та обмеження. Особливості біології як наукової дисципліни полягають у тому, що теоретичні узагальнення мають переважно емпіричний характер і багато в чому спираються на розуміння та віру, а не на доведеність та постійні підтвердження в природі. Майже кожне біологічне явище є унікальним (саме тому ми не можемо відтворити будь-які еволюційні події минулого або передбачити, що станеться в майбутньому), а тому може потребувати свого особистого правила, яке наука також не в змозі буде перевірити.