Основи еволюції - Корж О.П. - 2006
Частина I. МІКРОЕВОЛЮЦІЯ
Розділ 2. Популяція як елементарна еволюційна одиниця
2.2. Основні характеристики популяції
Однією з найважливіших характеристик популяції є її чисельність.
У більшості видів визначення меж окремого організму не вимагає особливих зусиль. Однак це стосується лише унітарних організмів, будова яких зумовлена генетично. Етапи розвитку, як і сама будова цих організмів, є цілком передбачуваними (за винятком суттєвих порушень): два вуха у зайця, шість ніг у комах, два крила у птаха тощо.
У модулярних організмів із зиготи розвивається певна одиниця будови (модуль), яка здатна давати початок новим модулям, що нагадують початковий. Ці організми майже завжди є розгалуженими і нерухливими. Складаються вони з певного набору основних елементів, кількість яких залежить переважно від взаємодії організму з довкіллям, а основною програмою їх розвитку є накопичення модулів, призначених для виконання певних функцій. Такі модулі можуть давати початок новим організмам, унаслідок чого з однієї зиготи утворюється кілька особин (загальна сукупність таких особин називається клоном). На певному етапі розвитку з'являються спеціалізовані репродуктивні модулі, які нових модулів здебільшого не утворюють, а дають початок новим зиготам.
Найбільш яскравими прикладами модулярних організмів є рослини, хоча трапляються й серед тварин (рис. 2.1). У більшості екосистем саме вони є організмами, що утворюють середовище (дерева в лісі, корали на коралових рифах тощо). Таким чином, стосовно модулярних організмів проблема полягає не лиіпе у вивченні кількості особин, а й у дослідженні чисельності та поширення модулів.
Загалом між модулярними та унітарними організмами існують певні відмінності: а) лише в модулярних організмів тіло може водночас складатися з молодих, активно функціонуючих та старих модулів (унітарні організми в певний момент можуть знаходитись лише на одній стадії розвитку); б) систематика модулярних організмів побудована переважно не за ознаками всього організму, а за його окремими модулями; в) взаємодія модулярних організмів із середовищем визначається їх загальною будовою, можливість «рухів» обмежена «переростанням» на нові місця або утворенням спеціалізованих модулів для розселення.

Рис. 2.1. Приклади модулярних організмів серед рослин і тварин (рис. О.В. Карнаухова)
а - рястка (Lcmna sp.) та гідра (Hydra sp.) з ростом організму розпадаються на частини; б - папороть (Dryopteris filix mas) та кампануларія (Campanullaria sp. типу Cnidariaкласу Hydrozoa) розростаються в горизонтальному напрямку; в - вівсяниця (Festuca octoflora) та мохуватка (Cryptosula sp.) складаються зі щільно прилягаючих модулів; г - дуб (Quercus sp.) та корал (Gordonia sp.) є -довгоіснуючими розгалуженими організмами
Виходячи з цього чисельність унітарних організмів можна визначити як:
Nn = Nt + В - D + С - Е,
де Nn - сучасна чисельність особин; Nt - попередня чисельність; В - кількість народжених за цей проміжок часу; D - кількість загиблих; С - іммігранти; Е - емігранти.
Стосовно модулярних організмів виникає необхідність у додатковій формулі:
Nn = Nt + В - D.
Кількість сучасних модулів «дорівнює» кількості попередніх модулів «плюс» кількість знову відроджених, «мінус» кількість відмерлих.
Розрізняють загальну чисельність особин, тобто загальну кількість особин на всіх стадіях розвитку, ефективну величину чисельності - кількість особин, що здатні розмножуватись, та мінімальну чисельність, тобто критичну величину, нижче якої популяція приречена на вимирання. Спроба визначення мінімальної життєздатної популяції (йдеться про чисельність, при якій популяція ще здатна існувати досить тривалий час), була безрезультатною, оскільки з'ясувалося, що єдиної кількості особин, яка б забезпечувала цю життєздатність відносно будь-якого організму, насправді не існує. У кожному окремому випадку необхідно підходити індивідуально, враховуючи видову належність організмів, місце, час та інші чинники.
Однією з найважливіших рис будь-якого таксону поряд з його морфологічними та екологічними характеристиками є ареал, який він займає.
Ареал - це частина земної поверхні (територія або акваторія), у межах якої поширений певний таксон і відбувається повний цикл розвитку його представників.
Популяція як частина виду також має свій певний ареал. Причому при його визначенні не враховується постійність мешкання особин та наповненість простору. Структура ареалів у різних організмів може бути досить складною, їх класифікація, запропонована Н.Ф. Реймерсом, наведена в схемі (рис. 2.2).
Ареали видів формувалися внаслідок взаємодії процесів еволюції самих тварин або рослин та зміни середовища їх мешкання в різні геологічні епохи. Тому за станом сучасного ареалу як окремих популяцій, так і певних таксонів у цілому можна скласти уявлення про процеси, що відбуваються з тією або іншою групою організмів.
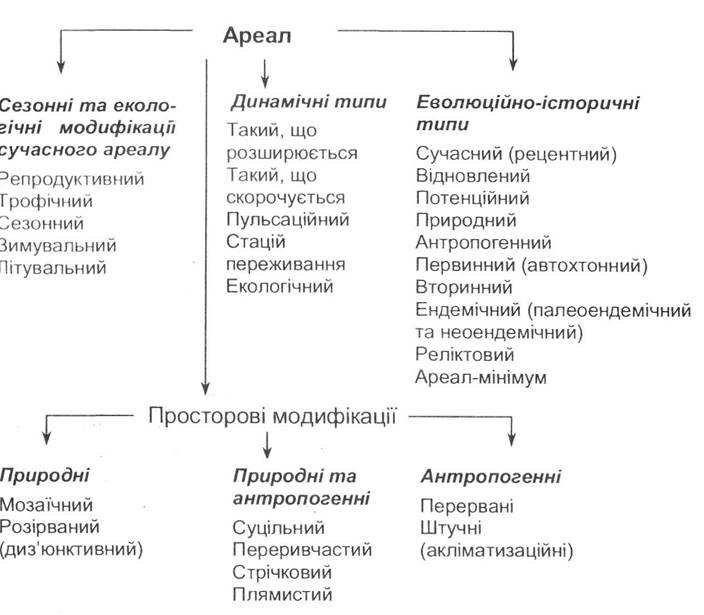
Рис. 2.2. Класифікація ареалів видів за Н.Ф. Реймерсом (1991)
Широтна складова ареалу є його основною географічною одиницею, оскільки свідчить про зональне поширення певних організмів. Вона визначається насамперед кліматичними, зокрема температурними, параметрами (часто їх розрізняють за зонами - арктична, бореальна, степова тощо). Висотна складова ареалу певного мірою аналогічна попередній, бо також визначається переважно за температурними умовами (альпійські, субальпійські та монтанні форми). Довготна складова визначається ступенем віддаленості від океану, що зумовлює характер клімату (аридний, або континентальний, та гумідний, або морський).
Ареали характеризуються різноманітністю, причому майже кожен з них унікальний. Форма та розміри ареалів залежать від цілого комплексу як сучасних, так і історичних причин. Однією з умов обмеження розмірів ареалу певної популяції може бути радіус репродуктивної активності особин, який часом сягає від кількох метрів (особливо це стосується рослин) до кількох тисяч кілометрів (водні та літаючі тварини).
Наслідком структури ареалу та чисельності популяції стає щільність особин.
Цей показник також досить важливий, оскільки за умови низької щільності здатні до розмноження особини можуть не знайти одна одну в межах відносно великого ареалу (подібна ситуація нині простежується в багатьох рідкісних та зникаючих видів).
Важливою характеристикою, пов'язаною з попередніми ознаками популяції, є мереживо ареалу виду. У природі завжди спостерігається скупчення особин в одних місцях і різке розрідження аж до повної їх відсутності в інших, що робить середовище схожим на мереживо. Це призводить до того, що межі популяції майже завжди є нечіткими. Найбільш яскраво таке явище простежується на прикладі плямистого ареалу (рис. 2.3).
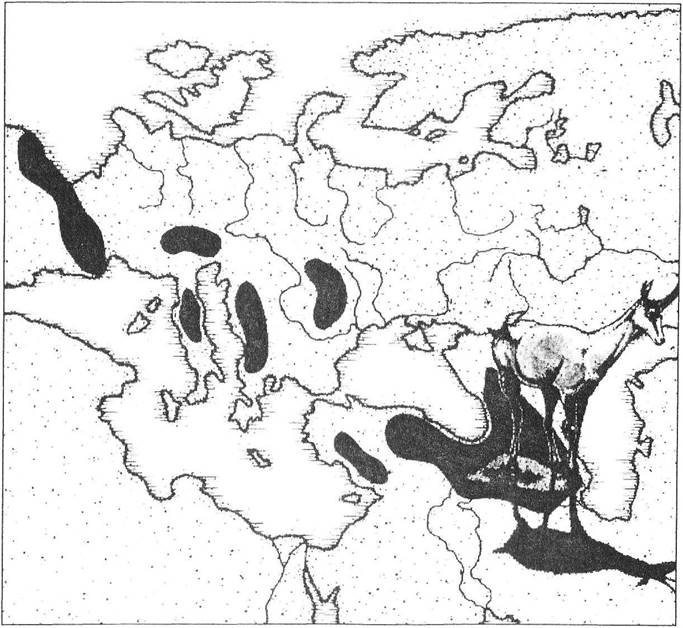
Рис. 2.3. Мозаїчний ареал на прикладі поширення сарни (Rupicapra rupicapra) в Європі (рис. О.В. Карнаухова)
Розміщення особин у просторі буває трьох видів, причому всі вони пов'язані між собою певними механізмами. Випадкове розміщення виникає тоді, коли будь-яке місце може бути зайняте з однаковою вірогідністю, а присутність одного організму ніяк не впливає на присутність інших. Регулярне, або рівномірне, розміщення формується в тому випадку, коли особини, що виявилися досить наближеними одна до одної, гинуть чи залишають популяцію. Групове розміщення створюється за умови приваблення особин популяції до конкретних ділянок місць існування.
При визначенні типу розміщення певних організмів слід враховувати масштаб, який використовується в конкретному випадку. Розміщення організмів будь-якого виду ареалом має переважно випадковий характер, однак у межах окремої популяції воно може стати груповим (кілька зграй вовків, мурашині колонії тощо). При розгляді ще дрібніших структур розміщення виявляється переважно рівномірним, що регламентується потребою в просторі для кожної особини (вовки всередині зграї, попелиця на листку тощо).
Життєві цикли як результуюча біологічних потенцій виду. Як зазначалося раніше, чисельність є результуючою кількох різноспрямованих процесів, з яких найбільш важливими є народжуваність та смертність. Саме вони, а також деякі інші біологічні особливості організмів складають специфіку їх життєвих циклів, які, у свою чергу, зумовлюють особливості динаміки популяцій.
Річний життєвий цикл звичайно триває близько 12 місяців, і особини вже до початку наступного сезону розмноження гинуть. Тому подібні покоління називають такими, що не перекриваються. Особини можуть брати участь у розмноженні лише один раз, після чого гинуть, або кілька разів, але не більше, ніж упродовж року. Прикладами таких життєвих циклів можуть бути однорічні трав'янисті рослини (деякі злаки, представники роду Veronica тощо), а також значна кількість переважно дрібних безхребетних тварин (деякі прямокрилі, метелики, перетинчастокрилі та ін.).
Цикл з багаторазовим розмноженням і перекриванням поколінь властивий організмам з порівняно великою тривалістю життя, але з розмноженням лише в певні сезони року. Прикладами подібних життєвих циклів можуть бути хребетні тварини з сезонним розмноженням, дерева помірної зони тощо.
До варіантів зазначених вище типів життєвих циклів слід віднести: дворічні організми, розмноження яких відбувається на другий рік їх існування; багаторічні організми з невизначеною тривалістю життя і одноразовим розмноженням, після чого вони гинуть; організми з безперервним багаторазовим і безперервним одноразовим розмноженням (останні, як правило, дають за рік кілька генерацій).
Особливості життєвих циклів, а також екологічні взаємовідносини організмів з довкіллям визначають динаміку чисельності популяцій. У певні сезони року вона може значно відрізнятись, що характерно переважно для дрібних організмів з високою репродуктивною здатністю, але й з великою смертністю. Різка зміна чисельності деяких видів пов'язана з наявністю кількох фаз розвитку, унаслідок чого аналізувати стан популяцій подібних видів слід лише з урахуванням стану особин, що знаходяться на всіх можливих фазах розвитку.
Причини різноманіття та формування конкретних життєвих циклів поки що залишаються загадковими і потребують подальших досліджень. Вважається, що розміри організмів є однією з передумов формування особливостей життєвих циклів. Більші за розмірами організми конкурентоспроможніші, краще зберігають постійний рівень функціонування в умовах коливання параметрів середовища, що сприяє підвищенню їх виживання. Із зростанням зазначених параметрів (у першу чергу - розмірів тіла) відповідно збільшується ймовірність постраждати від усіляких негараздів.
Формування певного життєвого циклу багато в чому залежить від динаміки народжуваності й смертності організмів. Найбільш класичними є три типи кривих виживання - опукла, яка стосується переважно К-стратегів (характеризується порівняно низькою репродуктивною здатністю та піклуванням про нащадків, що сприяє низькому рівню загибелі на перших етапах розвитку організмів); прямолінійна, при якій імовірність загибелі з віком не змінюється, та увігнута, властива r-стратегам, у яких найбільший відсоток загибелі припадає на перші етапи онтогенезу. Встановлено, що в чистому вигляді такі криві майже ніколи в природі не трапляються. У більшості видів така крива має вигляд ламаної лінії, що відповідає періодам з підвищеною загибеллю (кризові періоди), які чергуються з відносно сприятливими умовами існування і низькою загибеллю. Особливо така періодичність простежується у видів з кількома фазами розвитку життєвого циклу.
При вивченні багатьох унітарних організмів, крім народжуваності й смертності, слід враховувати наявність міграційних процесів, які в деяких випадках можуть спричинятися до більших змін у динаміці чисельності, порівняно з попередніми параметрами. Популяції багатьох організмів упродовж свого життя можуть здійснювати багаторазові переміщення з одного місця існування до інших з тривалістю міграційних циклів від кількох діб до місяців і навіть років. Досить часто розмежованість біотопів, у яких відбувається живлення тварин та їх відпочинок, змушує останніх до постійного добового пересування з одного місця до іншого.
Сезонні переселення можуть пов'язуватися зі зміщенням ресурсів відповідно до сезонів року з одних ділянок ареалу до інших. Зокрема, вертикальні міграції копитних у горах зумовлені пишним розвитком трав'янистої рослинності влітку на високогір'ях та більш м'яким кліматом взимку в долинах.
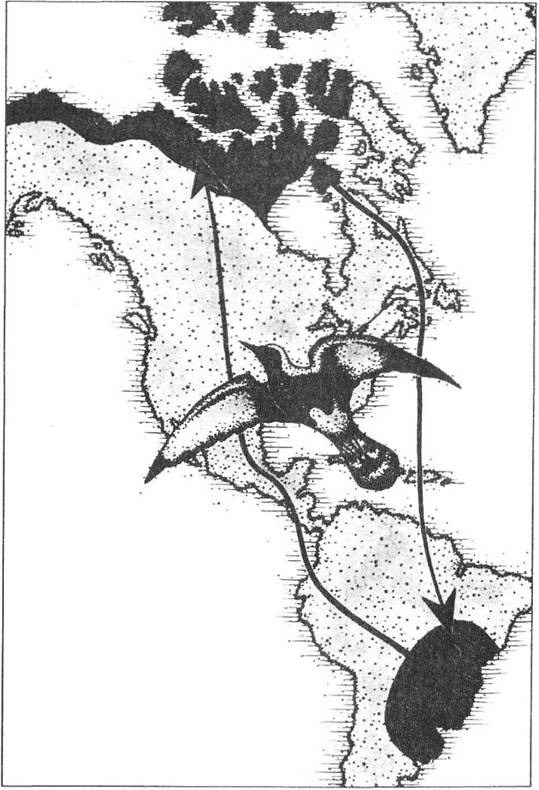
Рис. 2.4. Складний ареал і шляхи міграцій сивки бурокрилої (Pluvialis dominica) у Південній та Північній Америці (рис. О.В. Карнаухова)
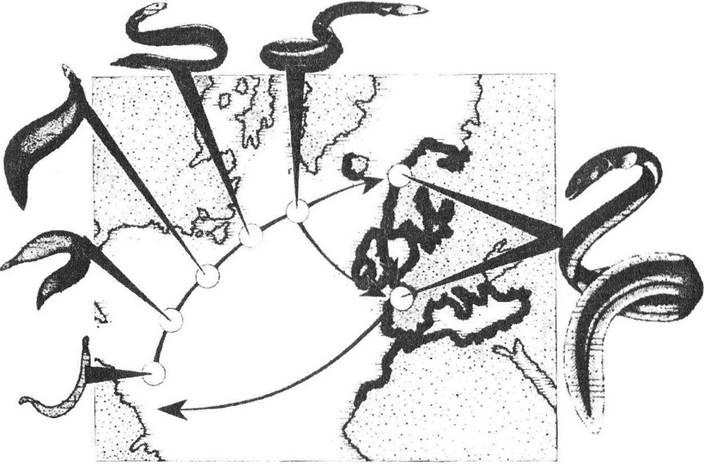
Рис. 2.5. Схема міграцій вугра європейського (Anguilla anguilla) до місць нагулу в Європі та розмноження в Саргасовому морі
(рис. О.В. Карнаухова)
Найбільш цікавими є міграції на значні відстані (рис. 2.4, 2.5). Зміст їх полягає переважно в переміщенні особин з одної віддаленої області до іншої, у кожній з яких є надлишок їжі, але лише впродовж певного сезону. Оскільки в інший час умови для існування на тій самій території стають вкрай неприйнятними, то осіле перебування популяцій такої самої чисельності в межах лише однієї території стає майже неможливим. Відповідні втрати як популяції в цілому через підвищену загибель під час міграцій, так і окремих особин через їх виснаження компенсуються наявністю значної кількості «вільних» ресурсів, завдяки чому і відбулося еволюційне закріплення подібних міграцій.
Вікова та статева структура популяції майже завжди знаходиться в динаміці та є унікальною.
Вікова структура популяцій визначається співвідношенням різних вікових груп організмів, що залежить від особливостей
життєвого циклу певного виду та конкретних умов існування. Крім календарного віку (час існування організмів), велике значення для популяції має біологічний вік, який свідчить про стадії онтогенезу, на яких перебувають ті чи інші групи особин. Віковий склад популяції характеризує її властивості, які відображають онтогенетичний стан останньої як цілісної системи.
Більш просту вікову будову має популяція, яка складається з особин майже одного віку. Вона властива деяким однорічним рослинам і тваринам. Наочним прикладом її можуть бути штучні популяції однорічних культур (кукурудза, горох, соняшник тощо) в агроценозах. У природних системах навіть у цьому випадку межі календарного віку двох суміжних вікових етапів можуть дещо перекриватися внаслідок індивідуальної мінливості в особливостях розгортання онтогенетичних змін. Так, навесні в багатьох гризунів, яким властивий однорічний цикл (велика піщанка - Rhombomys opimus та ін.), особини, що перезимували, дають два приплоди, а в деяких випадках і три. До того ж окремі самиці першого приплоду поточного року також можуть брати участь в останньому розмноженні. Після цього дорослі тварини гинуть, і на наступну весну залишаються лише особини попереднього року народження.
Таким чином, відносно гомогенні, на перший погляд, популяції складаються з кількох когорт (послідовні виводки однієї когорти батьків) і навіть генерацій (поколінь). Було з'ясовано, що сезонні вікові когорти можуть мати для популяції різне значення. Подібні вікові відмінності позначаються не лише на загальних параметрах росту й розвитку, але й на особливостях, що впливають на біологічну активність та місце популяції в біоценозі.
Тварини весняної та ранньолітньої когорт відзначаються підвищеним рівнем метаболізму і, як наслідок, незначною тривалістю життя - переважна більшість їх не доживає до наступної весни. Пізньолітні та осінні когорти мають протилежні властивості, тобто повільний ріст, але більш тривале життя. Тварини цих когорт забезпечують успішне долання популяцією скрутного за екологічними умовами зимового періоду, зберігаючи здатність до розмноження навесні. Тварини ранньовесняних когорт (нащадки попередньої групи) відрізняються високою швидкістю статевого дозрівання і забезпечують швидке відновлення чисельності популяції.
Природні популяції можуть складатися з особин кількох послідовних років народження. Так, вікову структуру популяції крота звичайного (Таіра еигораеа) утворюють цьоголітки (60-70%) та інші особини - віком від 1 до 5 років. В ондатри (Ondatra zibethica) переважну частину популяції становлять тварини до одного року, але трапляються дворічні і навіть трирічні особини.
Значно ускладнюється вікова структура популяції в тварин, особини яких різняться не лише за календарним, але й за біологічним віком. Так, до популяції ведмедів (Ursus arctos) можуть входити ведмежата-цьоголітки, лончаки (яким більше року і які йдуть від матері), поодинокі ведмеді, що не беруть участі в розмноженні, та особини-плідники. Крім того, у ній часом перебувають і старі тварини, які участі в розмноженні не беруть через неконкурентоспроможність із молодшими особинами, але в природних популяціях подібна категорія трапляється відносно рідко.
Слід враховувати, що в природі співвідношення різних когорт і генерацій за сезонами року завжди змінюється. Цей показник залежить від участі в розмноженні особин різного віку і відповідних передумов. У деревних рослин з їх порівняно тривалим життям структура популяції може бути складнішою, що зумовлюється взаємним заплідненням старих і молодих особин. Вікова структура популяції є показником репродуктивних властивостей, здатності до захоплення простору, процесів саморегулювання тощо.
Для популяцій бісексуальних видів, крім співвідношення окремих вікових груп, велике значення має ще й статева структура. Наявність статі, тобто розвиток усередині виду двох типів особин, здатних до схрещування, є загальною рисою майже всіх еукаріотичних організмів. Формування статей пов'язане з анізогамією (різні розміри статевих клітин), що добре простежується на формах з оогамією (утворення нерухливої яйцеклітини і дрібних рухливих сперматозоїдів). Проте воно необов'язкове, оскільки існують і гермафродитні види, які мають певні переваги: усі особини популяції в цьому випадку здатні виводити молодь, а поодинокі особини можуть також служити для розширення ареалу відповідного виду. Стосовно різностатевих організмів, то далеко не всі з них беруть участь у розмноженні. Відомий приклад, коли в морських слонів 4% самців запліднювали 88% самиць. Отже, деякі самці за своє життя не отримують навіть можливості реалізувати свій репродуктивний потенціал, хоча використовують ресурси популяції так само, як і інші, більш спритні тварини. Вважається, що наявність різностатевих особин (у першу чергу це стосується самців, які інколи розглядаються як не зовсім потрібний надлишок) необхідна для забезпечення нових комбінацій генів і підтримання різнорідності в популяції.
Співвідношення статей має значення для реалізації репродуктивного потенціалу та саморегуляції (самопідтримання) популяції. Крім цього, фізіологічні та екологічні відмінності самців і самиць сприяють зменшенню внутрішньопопуляційної конкуренції, бо мають дещо різні потреби. Таким чином, підтримання біологічної різноманітності в популяції також може вважатися однією з причин формування різностатевості.
Найбільш чітко статева структура популяції простежується на членистоногих та хребетних тваринах, яким притаманні не лише кількісні відмінності між статями, а й просторово-функціональні. Відповідна динаміка багато в чому пов'язана з віковою структурою, у зв'язку з чим розрізняють первинне, вторинне й третинне співвідношення статей.
Первинне співвідношення визначається суто генетичними механізмами, що спираються на різноякісність статевих хромосом. У ссавців, як і в більшості інших тварин, самці й самиці мають різний набір статевих хромосом (XY та XX). Цей механізм створює передумови для рівного співвідношення статей у нащадків, яке й приймається за первинне. Проте навіть під час запліднення первинне співвідношення статей може порушуватись через неоднакову здатність сперматозоїдів до запліднення або через вибірність яйцеклітин. До цього слід додати можливий вплив середовища, що також може позначитись на розвитку певних зигот або на інших етапах формування статі майбутньої особини.
Вторинне співвідношення — кількість самців та самиць серед новонароджених особин. Воно може значно відрізнятись від первинного через неоднаковий рівень загибелі ембріонів різної статі.
Третинне співвідношення статей властиве дорослим тваринам що розмножуються, і складається внаслідок неоднакової кількості втрат особин певної статі в ході їх онтогенезу. Саме воно може свідчити про репродуктивний потенціал популяції й значно відрізняється у представників різних систематичних груп.
Найбільш докладно демографічні процеси вивчені в людини, що пов'язано з практичною необхідністю. Вікова структура популяції розвинених країн з низькою народжуваністю та смертністю суттєво відрізняється в державах, що розвиваються (висока народжуваність при відносно високій смертності). Статеве співвідношення залежить від багатьох факторів, але в цілому воно складається таким чином: незначне переважання хлопчиків у вторинному співвідношенні, рівний склад на час досягнення повноліття і подальше переважання жінок.
Структура популяції має певне значення для її подальшої долі.
Усі популяції безперервно змінюються, оскільки нові організми народжуються або іммігрують, а попередні гинуть чи емігрують. Незважаючи на це, флуктуації розмірів популяцій ніколи не бувають безмежними: вони не можуть нескінченно зростати, але й вимирають відносно рідко.
Питання про механізми регулювання чисельності популяції спочатку розглядали як дихотомічну альтернативу — дію внутрішніх чи зовнішніх чинників. Сьогодні вже зрозуміло, що і структура популяції, і її динаміка залежать від цілого комплексу умов, у яких опинилась та чи інша популяція. Жодні екологічні фактори не можуть впливати на будь-який вид самостійно, незалежно один від одного, унаслідок чого і виникає необхідність використання в аналізі популяційних процесів інтегральних показників.
При вивченні еволюційного процесу важливе значення має дослідження генофонду популяції (сукупність генотипів усіх особин). Безпосередньо ми можемо спостерігати лише фенотипи (сукупність зовнішніх ознак), а не окремі гени чи навіть генотипи (набір генів однієї особини). Мінливість генофонду може бути описана частотами генів або генотипів. Природним популяціям притаманна генетична мінливість, причому вона значно перевищує морфологічну різноманітність, яку ми спостерігаємо.
Математичні розрахунки свідчать, що навіть два організми (крім монозиготних близнюків або клонів) не можуть бути тотожними один одному, а також жодному з попередніх і майбутніх організмів. Таким чином, виникає генетична гетерогенність популяції. Її підтриманню в природі сприяє поява мутацій, комбінативна мінливість і кросинговер, які ще більше урізноманітнюють її за рахунок нових ознак та формування їх нових сполучень.
Незважаючи на відмінності окремих особин, тобто їх гетерогенність, популяція характеризується генетичною єдністю. Саме вона є мінімальною за розмірами генетичною одиницею, яка здатна до самостійного існування впродовж значної кількості поколінь. Завдяки панміксії (вільному схрещуванню) складна генетична структура популяції знаходиться в стані динамічної рівноваги. Через відсутність необхідної кількості статевих партнерів для підтримання генетичного різноманіття ні окрема особина, ні сім'я, ні інші, дрібніші за популяцію, угруповання особин таких властивостей не мають.
Разом з тим не завжди навіть усередині популяції панміксія буває повною. На вибір статевого партнера може впливати його генотип (таке явище називають асортативним схрещуванням). Наприклад, кількість шлюбів, які укладаються між двома білими або двома неграми в Америці, перевищує кількість змішаних пар, що свідчить про певну вибірність статевого партнера за кольором шкіри. Проте це не означає, що подібна вибірковість стосується й інших ознак, наприклад групи крові. Асортативне схрещування в природі притаманне не лише людині - його крайньою формою можна вважати самозапліднення.
До особливостей певної популяції відносять її екологічну нішу. Екологічна ніша - це сукупність усіх факторів середовища, у межах яких можливе існування виду в природі (або екосистемі), та його середовищеутворююча діяльність. Іншими словами, це місце виду переважно в біоценозі та його підрозділах, причому не лише в розумінні просторового розташування, а й функціональної ролі.
Враховуючи те що в жодного виду не існує двох подібних популяцій, слід визнати, що місце цих популяцій у відповідних біогеоценозах, а, як наслідок, і відповідна екологічна ніша, є унікальними. Теоретично можна припустити наявність «вільних» екологічних ніш у певному біогеоценозі - недовикористаних ресурсів, які активно використовуються в інших подібних екосистемах. В реальності кожна екологічна ніша виникає разом з формою, яка її займає.
Наведений вище матеріал свідчить про унікальність кожної популяції.
Популяції будь-якого виду відрізняються одна від одної статистично майже за всіма показниками. Це стосується чисельності та її динаміки, статевого й вікового складу, а головне - генетичної структури. Відмінним у популяціях є також кількісне співвідношення різних алелей і частот виявлення того чи іншого фенотипу.
На рівні популяції індивідуальна різноякісність доповнюється відмінностями у складі та особливостях функціонування окремих внутрішньопопуляційних угруповань. Так, у копитних поряд із двостатевими угрупованнями в популяції існують окремі стада самців-холостяків, поодиноких особин, групи лише самиць та нестабільного складу. У популяціях кашалотів (Physeter catodon) описано понад шість типів внутрішньопопуляційних угруповань. Слід зазначити, що подібні угруповання можуть відігравати різну роль у житті популяції. Зокрема, холостяцькі стада (складаються переважно з молодих тварин) виводять молодь з-під пригнічуючої дії домінантів, що дозволяє їм нормально розвиватись, доки вони не стануть здатними брати участь у розмноженні.
Навіть у видів з обмеженим ареалом окремі популяції мешкають в умовах різного рельєфу, рослинності, мікроклімату тощо. Усе це призводить до появи відмінностей морфобіологічного типу у тварин відповідних популяцій. Усередині однієї популяції, навпаки, особинам властива схожість морфобіологічного типу та єдність ритму життєвих відправлень, що пояснюється загальною спрямованістю пристосувань індивідів у схожих умовах. В еволюційному процесі популяція виступає як екологічна, морфофізіологічна і генетична єдність, яку визнають елементарною одиницею, у межах якої може відбуватись еволюція. Лише вона, а не окрема особина чи група особин допопуляційного рівня, має свою еволюційну долю.
При цьому виникає питання: чи не можна визнати за популяцію кілька особин-засновників, які в подальшому дають початок справжній популяції? Прикладами можуть бути кролики Австралії, ондатра й колорадський жук Європи, більшість острівних форм. Проте ця невеличка група не може дорівнювати популяції, бо вона не має необхідних характеристик, і до часу формування справжньої популяції може загинути не одна подібна мікропопуляція. Так, у Підмосков'ї у 60-70-х роках XX століття створили штучну популяцію мисливського фазана, яка кілька років існувала в природі в кількості близько 30 особин. У зимовий період її підгодовували, але, незважаючи на це, в одну з особливо суворих зим вона повністю вимерла. Таким чином, долю кожної мікропопуляції можна зрозуміти лише з часом.
Популяція до певної міри має ті самі властивості, що й окрема особина — може виникати, рости, диференціюватися тощо. Але, на відміну від особини, вона може існувати в часі необмежено при більш-менш сприятливих умовах. Властивості популяції визначаються в першу чергу властивостями особин, що до неї входять, тобто залежать від їх віку, статі, фізіологічного стану тощо. Однак, крім цього, важливими характеристиками стають просторова структура популяції, її динаміка і навіть відносини особин одна з одною. Тому популяції звичайно виступають як єдине екологічне та генетичне ціле.
Визнання популяції елементарною еволюційною одиницею свідчить про те, що всередині неї й відбуваються процеси, які зумовлюють еволюційні зміни. Біологічна еволюція - це процес накопичення змін в організмах і збільшення їх різноманіття. Підґрунтям для її здійснення стають генетичні, тобто спадкові зміни. Тому на генетичному рівні еволюцію можна уявити як накопичення змін у генетичній структурі популяції.
Елементарне еволюційне явище — це тривала спрямована зміна генотипного складу популяції.
Елементарне еволюційне явище ще не можна вважати еволюцією, але саме з таких елементів і складається увесь еволюційний процес.
Думки вголос
Отже, популяцію визнано як елементарну еволюційну одиницю. Однак на практиці окреслити реальні межі будь-якої популяції досить складно, тому дослідження останньої ґрунтуються переважно на вивченні вибірок. Питання про те, якими вибірками користувались - з різних популяцій, чи з однієї, - вирішується шляхом використання методів математичного аналізу. Таким чином, уявлення про процеси, що відбуваються всередині елементарної еволюційної одиниці, багато в чому є наслідком екстраполяції, тобто формуються на рівні припущення. Таким чином, реальні події в популяції можуть значно відрізнятись від наших уявлень про їх перебіг. Саме тому польові дослідження стану природних популяцій не завжди дають очікуваний матеріал для осягнення мікроеволюційних механізмів.