Основи еволюції - Корж О.П. - 2006
Частина III.ШЛЯХИ РОЗВИТКУ ЖИТТЯ
Розділ 19. Основні напрямки розвитку живих організмів
19.4.Виникнення насінини як етап еволюції вищих рослин
Риніофіти дали початок плавуноподібним, хвощеподібним та папоротеподібним, які виявилися краще пристосованими до наземного існування. Ці рослини відрізнялися значними еволюційними надбаннями порівняно зі своїми предковими формами: у них виникає типова судинна система, а в папоротей - навіть листя та коріння. Слабким їх місцем слід вважати збереження способу
розмноження, яке вимагало наявності водного середовища, бо, як і в риніофітів, чоловічі статеві клітини для запліднення повинні були доплисти до архегоніїв. Чергування життєвих поколінь (спорофіт — гаметофіт) у них також зберігається, хоча останній підпадає суттєвій редукції (так званий заросток).
З початком посухи в пермському періоді, а потім і в тріасі (близько 300 млн років тому), починається масове вимирання рослин, запліднення в яких потребувало водного середовища. У цей час на арену еволюції виходять насінні рослини (рис. 19.7), у яких цей процес здійснюється без краплинно-рідинного середовища (ароморфна риса).
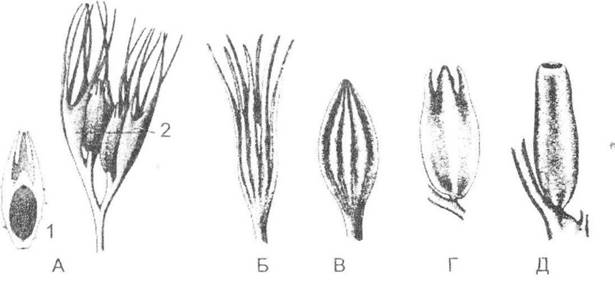
Рис. 19.7. Насіннєподібні структури палеозойських рослин (з П. Рейвна, Р. Еверта та С. Айкхорна, 1990):
А - Archeosperma arnoldii; Б - Genomosperma kidstonii; В - G. lateens; Г - Euristoma angulare; Д - Stamnostoma huttonense;
1 - розташування мегаспори в насінині; 2 - купули (бокалоподібні утворення, що охоплюють мегаспорангії). Простежується поступова заміна пальцеподібних виростів (Б) на інтегументні лопасті, які повністю зростаються (Д)
Усі насінні рослини, як і деякі папороті, є різноспоровими. Вони мають спори двох типів: мікроспори (чоловічі) та мегаспори (жіночі), які дають початок чоловічому та жіночому гаметофітам відповідно. Але в насінних рослин, на відміну від безнасінних, єдина зріла мегаспора не залишає мегаспорангія - там відбувається розвиток жіночого гаметофіта та запліднення.
Оскільки насінина має захисну оболонку, усередині якої знаходиться зародок та поживні речовини для його розвитку, вона стає ефективним способом розмноження порівняно зі спорами.
Внутрішнє запліднення, розвиток зародка всередині насінного зачатка, а також поява дуже ефективної одиниці розмноження - насінини — є головними біологічними набутками насій них рослин, які й дали можливість останнім краще пристосовуватись до умов суходолу й зайняти значно важливіше місце в рослинному світі.
Голонасінні рослини суттєво відрізняються від спорових як розвитком гаметофіта, так і самим процесом запліднення. Жіночий гаметофіт в усіх голонасінних рослин розвивається всередині мегаспорангія і ніде не контактує з повітряним середовищем. Доступ до нього здійснюється лише через мікропіле (пилковхід), що створює найбільш сприятливі умови для захисту жіночого гаметофіту від висихання. Як наслідок, відбувається поступова редукція й спрощення жіночого гаметофіту з архегонієм та починається раннє формування яйцеклітини.
Чоловічий гаметофіт також спрощується, а його розвиток значно прискорюється. На відміну від папоротей у голонасін них перший поділ ядра мікроспори відбувається ще всередині мікроспорангія. Свого остаточного розвитку чоловічий гаметофіт набуває на мегаспорангії, паразитуючи на останньому. Він уже зовсім позбавлений антеридіїв.
Виникненню голонасінних рослин передувало кілька важливих еволюційних подій, які сталися переважно в девоні. У сфері репродукції — це формування різноспоровості (різноспорові форми відомі ще з середнього девону - близько 400 млн років тому). Вегетативна частина рослин також набула деяких змін: приблизно в цей час виникає камбій, який надає рослинам можливість вторинного росту, а згодом з'являються й деревні форми.
Оскільки перші голонасінні виникли близько 370 млн років тому, їх не можна виводити зі справжніх папоротей (існували водночас, а деякі з останніх за своєю організацією перевищують примітивні голонасінні). Таким чином, безпосередніх предків голонасінних слід шукати серед примітивних папоротеподібних - різноспорових деревних форм, яким був властивий вторинний ріст.