Основи еволюції - Корж О.П. - 2006
Частина III.ШЛЯХИ РОЗВИТКУ ЖИТТЯ
Розділ 20. Основні етапи розвитку багатоклітинних тварин
20.4.Розвиток порожнини тіла
Первинна порожнина тіла виникає в круглих черв'яків: мезенхіма в них зникає, а простір між кишкою та м'язошкірним мішком (щілини між внутрішніми органами) займає первинна порожнина тіла. Ця порожнина заповнена рідиною, значний тиск якої відіграє роль скелета. Оскільки кільцеві м'язи відсутні, тварини можуть здійснювати лише змієподібні рухи.
Первиннопорожнинним черв'якам властива постійність загального плану будови тіла та клітинного складу. Епідермальні клітини об'єднані в синцитій, під ним розташований один шар поздовжніх м'язових волокон. Тіло не сегментоване, дихальна та кровоносна системи відсутні, у травній системі з'являються задня кишка та анальний отвір. Видільна система або відсутня, або протонефридіального типу; може бути представлена шкірними залозами. Нервова система побудована, як і в плоских черв'яків (ортогон). На відміну від плоских власне круглі черв'яки є первинно різностатевими тваринами. Незважаючи на досить просту організацію, первиннопорожнинні є дуже поширеними організмами, які відносно добре пристосовані до найрізноманітніших умов існування. Чимало з них стають паразитичними формами, ускладнюючи свій життєвий цикл, інколи набуваючи гермафродитизму та чергування поколінь.
Вважається, що філогенетично цей тип пов'язаний з плоскими черв'яками, про що свідчить наявність окремих рис останніх у різних класах первиннопорожнинних. Нервова система нематод майже така сама, як і в турбелярій. Глотка в примітивних класів первиннопорожнинних має однакову будову з глоткою прямокишкових турбелярій. Розташування рота на передньому кінці тіла (як в усіх первиннопорожнинних) характерна для представників ряду WiabdocoeLa. Статева система не має суттєвих відмінностей у своїй будові, за винятком гермафродитизму. Іншою спільною ознакою круглих та плоских черв'яків слід вважати наявність у примітивних коловерток та черевовійчастих ділянок тіла, вкритих війками.
Целом — вторинна порожнина тіла, яка являє собою переважно міхури, заповнені всередині целомічною рідиною. З розвитком целому первинна порожнина тіла підпадає редукції і витісняється останнім.
Існує кілька теорій походження целому:
1. Схізоцельна теорія: целом анелід є вдосконаленням схізоцеля первиннопорожнинних тварин. Ця теорія не пояснює, чому в ембріогенезі у багатьох тварин целом формується за рахунок стінок кишечника.
2. Міоцельна теорія (різновид попередньої): целом виникає як порожнина тіла в м'язовому зачатку, який потім заповнюється рідиною. Отже, згідно з цією теорією, спочатку целом виконував опорну функцію (інші функції вторинної порожнини тіла не пояснюються).
3. Гоноцельна теорія припускає, що целом виникає зі статевих залоз нижчих черв'яків: порожнина кожного целомічного
міхура відповідає порожнині гонади турбелярій або немертин. Таким чином, перітонеальний епітелій виводиться зі стінок статевих залоз (далі робиться припущення, що частина статевих клітин перетворюється на соматичні, це є, на наш погляд, досить сумнівним).
4. Ентероцельна теорія: целом бере свій початок від гастроваскулярної порожнини кишковопорожнинних та реброплавів, яких можна вважати віддаленими предками целомічних тварин. Саме ця теорія вважається найбільш прийнятною, оскільки пояснює статеву функцію целому (вбирає в себе гоноцельну теорію) та підтверджується ходом розвитку целому в деяких тварин, зокрема вторинноротих.
В анелід уперше з'являється справжня метамерія тіла, яка простежується в їх зовнішній та внутрішній будові, хоча такі риси мали й первинні війчасті черв'яки. Сегментація тіла первинних анелід носить гомономний характер, а в спеціалізованих форм проявляється гетерономія.
Походження анелід деякі вчені (зокрема, В.Н. Беклемішев) пов'язують з реброплавами, спираючись на спільність деяких етапів їх розвитку. Існують і інші теорії - «турбелярна», «немертинова» тощо.
Неспеціалізовані форми поліхет характеризуються процесом цефалізації: кілька перших сегментів утворюють головний відділ. Однією з ароморфних рис анелід слід вважати виникнення дволопасних виростів - параподій, які виконують функцію руху та здійснюють у деяких форм газообмін (утворюють зябра). У цих тварин виникає також кровоносна система, яка ще не має сформованого серця і є первинно замкненою. У цілому цим тваринам притаманний процес олігомеризації органів та структур, який найглибше простежується в п'явок.
Найчисленнішим за видовим складом, причому не лише з-поміж тварин, є тип членистоногих (Arthropoda), який нараховує понад 1,5 млн видів (теоретично припускається, що реальна кількість видів перевищує кілька мільйонів). Його представники опанували всі середовища існування на Землі, і в кожному класі спостерігається широка адаптивна радіація. Філогенетично артроподи виводяться з примітивних анелід. Але в них є й кілька суттєвих ароморфних рис.
Бічні придатки тіла анелід (нерозчленовані параподії) в артропод перетворюються на членисті кінцівки - артроподі!, які являють собою кілька трубчастих члеників, з'єднаних суглобами. Вони можуть перетворюватися на сенсорні органи (антени), частини ротового апарата (мандибули, максили, хеліцери, педі-
пальпи тощо), локомоторні кінцівки (плавальні та ходильні) або копулятивні апарати. Саме такі кінцівки уможливлюють високу адаптаційну здатність артропод.
Видільна система цих тварин складається з перетворених целомодуктів (коксальні залози) або мальпігієвих судин. Це дозволяє представникам зазначеного типу значно ефективніше займати різні екологічні ніші.
Утворення кінцівок, побудованих за принципом важелів, та значне підвищення загального рівня організації артропод стає можливим саме завдяки формуванню зовнішнього скелета - кутикули. Особливу роль це утворення починає відігравати після переходу окремих груп, зокрема багатоніжок та комах, до наземного способу існування. У цьому разі гостро постає проблема збереження організмом вологи. Тому в кутикули виникає новий зовнішній дуже тонкий шар, який складається з водонепроникних воскоподібних та жироподібних речовин.
У більшості артропод триває розвиток процесів цефалізації - тагма голови складається з акрона та чотирьох (дехто вважає з п'яти) сегментів. Прогресивний розвиток нервової системи дозволяє членистоногим перейти на принципово новий рівень розвитку - соціальний (спостерігається в суспільних комах).
Наведені та інші ознаки піднесли функціональну організацію членистоногих так високо, що за видовим різноманіттям вони значно перевищили всі інші групи тварин і утворили значну кількість спеціалізованих форм. Але щільний зовнішній скелет і велика швидкість пересування значною мірою обмежують загальні розміри артропод, найбільшими представниками яких були ракоскорпіони завдовжки близько 1,8 м.
Походження найчисленнішого типу тварин багато в чому залишається загадкою. Більшість фахівців сходиться на тому, що предковою групою для артропод слід вважати якихось примітивних поліхет (рис. 20.2), але щодо детального розгортання подій можна навести принаймні два сценарії.
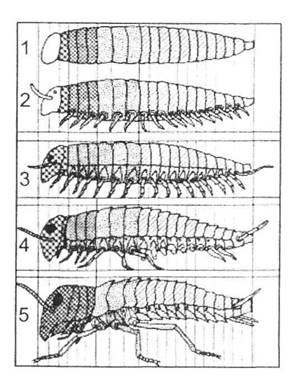
Рис. 20.2. Схема еволюції членистоногих від черв'якоподібних предків до комах за гіпотезою Р. Снодграса (за М.С. Гіляровим, 1984):
1 - членистий докембрійський черв'як без придатків; 2 - багатоніжкоподібна форма, схожа з ембріоном оніхофор, багатоніжок та комах; 3 - багатоніжкоподібна форма; 4 - перехідна форма від багатоніжок до нижчих комах; 5 - комахи
За одним з них, морські поліхети вийшли на суходіл, через що параподії пересунулися на черевну частину тіла й перетворилися на нечленисті ніжки (подібні утворення властиві Onychophora). Подальше пристосування до суходільних умов призвело до формування членистих кінцівок, ротового апарата й інших характеристик артропод - так з'явилися багатоніжки (Myriapoda), а далі й комахи (Insecta). У цьому випадку інші членистоногі (Trilobitomorpha, Crustacea, Chelicerata) походять від анелід незалежно від попередніх груп.
Згідно з іншим сценарієм, поліхети дають початок трилобіто- подібним, кінцівки яких розчленувалися не через наземний спосіб життя, а завдяки бентосному мешканню. Подібна структура кінцівок збереглася в їх нащадків - ракоподібних, а після виходу на суходіл - й у багатоніжок та комах. Проте існують свідчення про незалежне походження в деяких представників артропод трахейної системи, формування кінцівок, процесу цефалізації та деяких інших ознак. Тому в членистоногих пропонується розрізняти принаймні дві філогенетично незалежні гілки - Mandibulata nаmandibulata. У цілому можна зазначити, що особливості індивідуального розвитку членистоногих навіть у невеликих систематичних угрупованнях (особливо це стосується комах) набувають такого різноманіття, що не слід очікувати швидкого вирішення всіх проблем їх філогенетичних зв'язків.
До високорозвинених безхребетних також можна віднести один із поширених типів тварин - молюски (Mollusca). За однією з гіпотез, вони походять від анелід, що підтверджує знахідка молюска неопіліни (Neopilina galatheae та інші види), в якого присутні примітивні риси організації, що пов'язують їх з анелідами, а саме: добре розвинений целом, збереження сегментації в розміщенні нефридіїв, зябер, м'язів тощо. Потім ці структури безслідно зникають у молюсків: целом у них підпадає редукції (зберігається лише навколо серця та порожнини гонад), сегментація тіла зазнає повної редукції. Найбільш виразною ознакою всіх молюсків стає формування особливого органа — ноги, яка сприяє повзанню, плаванню тощо (у головоногих вона поділяється на кілька щупальців).
Предкова група молюсків, очевидно, відрізнялася відсутністю справжньої раковини (виникли ще у вендський період), невисоким тілом та більш активним способом життя порівняно з неопіліною. Від цього гіпотетичного предка дуже рано відокремилися дві головні гілки молюсків - Amp/iineura і Conchifera.
Переважна частина молюсків існує у водному середовищі, проте легеневі черевоногі опанували суходіл, виробивши засоби захисту від висихання спеціальний слиз, що закриває вхід у черепашку. Найрозвиненіший головний мозок з-поміж усіх молюсків мають головоногі. За розвитком нервової системи та можливостями створення умовних рефлексів їх можна порівняти з хребетними тваринами.