Основи еволюції - Корж О.П. - 2006
Частина III.ШЛЯХИ РОЗВИТКУ ЖИТТЯ
Розділ 20. Основні етапи розвитку багатоклітинних тварин
20.5.Походження вторинноротих
Походження хордових тварин (найбільш високоорганізованого типу вторинноротих) і сьогодні породжує масу теорій та гіпотез. Вважається, що необхідно розробляти теорії, які б пояснювали походження всієї групи вторинноротих, а не лише окремих типів. Вирішення проблеми деякі вчені вбачають у виведенні їх від найпростіших Metazoa. Однак наявність спільних рис організації (целом, мезодерма, схожість майже всіх систем органів тощо) у первинно- та вторинноротих вказує на неможливість їх набуття шляхом конвергенції.
Кілька теорій виникнення хордових висувалося ще у XIX столітті. За однією з них, предками цієї групи вважалися анеліди, оскільки обом групам властива білатеральна симетрія, сегментація тіла, центральна нервова система у вигляді головного мозку та поздовжнього нервового стовбура, а також переважно активний спосіб життя. Проте в іншому ці тварини дуже несхожі між собою: сегментація хребетних обмежується частиною мезодерми, нервовий ланцюг анелід розташовується на вентральній частині тіла, а в хребетних - на дорсальній; анеліди не мають хорди та зябрових отворів. Для подолання низки протиріч прихильники цієї теорії висунули положення про перевертання тварини в дорсовентральному напрямку, унаслідок чого аналогічно змістилися й системи органів. Але уявити його реальність - річ також неймовірна.
Предками хребетних пропонували вважати також павукоподібних. Ця гіпотеза ґрунтувалася на морфологічній схожості панцира викопних евріптерид (належать до павукоподібних) з кістковим панциром деяких викопних риб. Однак павукоподібні мають також черевний нервовий ланцюг, тому їм також необхідно було «перекидатися», що суперечить попереднім припущенням, оскільки схожість між цими тваринами зберігається лише в дорсальних та вентральних сторін тіла, а при його перевертанні зникає. Додатковою проблемою є члениста структура кінцівок, які не могли перетворитися на плавці риб (необхідно припустити попереднє зникнення цих кінцівок). До того ж павукоподібні не мають хорди та зябрових щілин. Таким чином, спочатку представники цієї групи повинні були втратити всі свої характерні ознаки, а вже потім перетворитися на хордових. Для зняття деяких протиріч було запропоновано модифікацію цієї теорії. Припускалося, що травна система павукоподібних, яка має складну будову, дає початок нервовій системі хребетних, а травна система потім виникає окремо. Але не існує жодних підстав припускати таку зміну функцій.
На сьогодні найпоширенішими є дві точки зору щодо походження вторинноротих. Згідно з ними, ці тварини походять від різних предкових форм, унаслідок чого філогенетичне дерево будується майже протилежним чином. За однією з теорій (дотримуються іноземні фахівці) предок вторинноротих був сидячою донною формою і живився поживними частками, які захоплював за допомогою руклофофорів (рис. 20.3). Від нього в разі розвитку спеціалізованих органів виникають голкошкірі, які дають початок геміхордовим. Розвиток ознак хордових починається зі зміни способу живлення: захоплення їжі лофофорами замінюється механізмом відфільтровування через зяброві щілини (як бічну гілку на цьому шляху розглядають групу геміхордових).
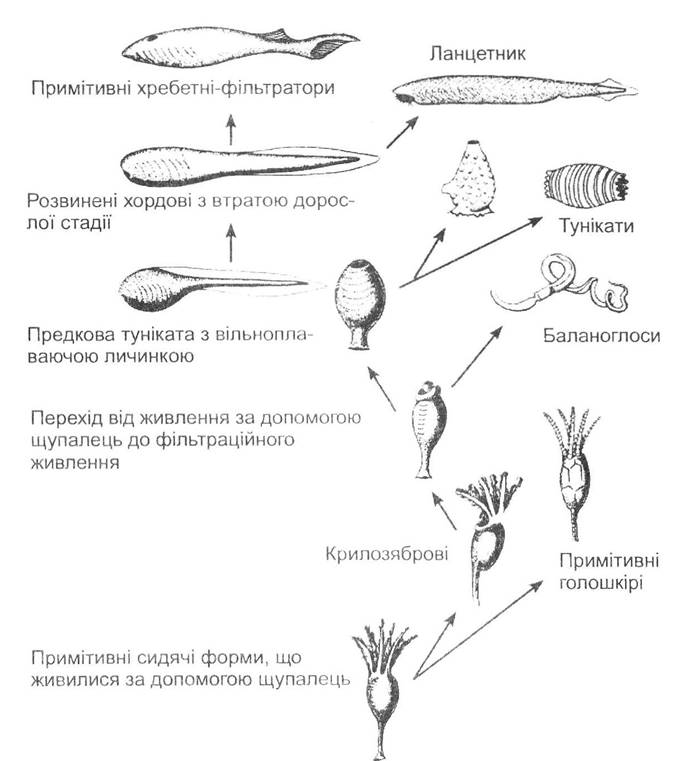
Рис. 20.3. Схема ймовірного шляху еволюції хребетних тварин (за А. Ромером та Т. Парсонсом, 1992)
Подальший розвиток та вдосконалення глотки як знаряддя для збирання їжі призвели до виникнення справжніх покривників (Tunicata), у яких доросла тварина являє собою спеціалізоване пристосування для фільтрування води. У той же час личинка цих сидячих тварин здатна вільно рухатись у пошуку місця для прикріплення й подальшого метаморфозу (виконує функцію розселення). З появою такої личинки виникає рухливий тип життя, що відкриває нові можливості для хордових тварин. Консервативні форми дали початок сучасним покривникам, а прогресивні, завдяки неотенії, перетворилися на тварин, схожих на ланцетника. Підтвердженням цієї думки вважається існування апендикулярій, які дійсно є неотенічними тваринами, але за своїм розвитком вони знаходяться вище, ніж ланцетник.
На нашу думку, ця теорія має певні суперечності. По-перше, голкошкірі тварини мають личинку з білатеральною симетрією, тому їх радіальна симетрія є вторинною. Це свідчить про те, що вони є дуже спеціалізованими. По-друге, покривники, на відміну від голкошкірих, не мають радіальної симетрії. По-третє, майже всім покривникам властива білатеральносиметрична личинка, яка за своїм розвитком стоїть значно вище порівняно з безчерепними. Таким чином, ця теорія не може вважатись вдалим поясненням виникнення вторинноротих тварин.
За іншою гіпотезою, яку розвивали О.М. Сєверцов, І.І. Шмальгаузен та інші, хребетні тварини виводяться з кишководихаючих, яскравим прикладом яких є баланоглос (Balanoglossus).
Найважливішою рисою, яка пов'язує цю тварину з хордовими, є її дихальний апарат (у глотці знаходяться кілька зябрових щілин, що підтримуються еластичним скелетом у вигляді решітки). Крім цього, кишечник має вигляд практично прямої трубки, кровоносна система складається переважно зі спинної та черевної судин, нервова система трохи схожа з хребетними, є також утворення, яке можна вважати зачатком хорди. У той же час деякі ознаки поєднують цю групу з кільчастими черв'яками (рис. 20.4).
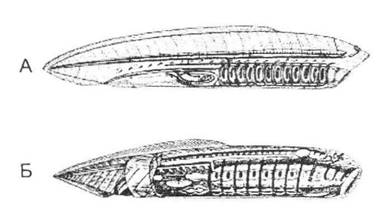
Рис. 20.4. Гіпотетичні предкові форми хребетних тварин (за О.М. Сєверцовим, 1945):
А - гіпотетичний предок хордових; Б - первинна черепна хребетна тварина
Оскільки первинно- та вториннороті тварини розділилися в ході еволюції досить рано, їх спільними предками слід вважати примітивних черв'яків на зразок турбелярій. Докладніше про безпосередніх предків поки що нічого не відомо. Кишководихаючі, голкошкірі та хордові відокремились від спільного стовбура, при цьому перші практично не змінилися в ході еволюції й максимально наближені до предкових форм вторинноротих.
Розвиток голкошкірих переважно пов'язаний з їх спеціалізацією (обмежена рухливість), що призводить до набуття радіальної симетрії, втрати целому тощо. Перетворення білатеральної личинки на дорослу форму з радіальною симетрією більше не характерне жодній із груп тварин. Переважна частина голкошкірих є рухливими, поверненими оральною частиною тіла до ґрунту тваринами. Але морські лілії є сидячими організмами (кріпляться до субстрату стебельцем або вусиками), їх ротовий отвір розташований на поверхні чашечки. Вважається, що спочатку всі голкошкірі живилися планктоном та іншими дрібними частками, це й зумовило їх радіальну симетрію та малу рухливість. Пізніше деякі стали хижаками й почали вести більш рухливий спосіб життя.
Жодна з сучасних форм не може вважатися безпосереднім предком сучасних тварин, а викопну форму, яка б могла дати початок хордовим, поки що не знайдено.
Якщо ж вдатися до припущення, то гіпотетичним предком хордових слід вважати представників безчерепних, які за своєю організацією наближаються до сучасного ланцетника. Саме вони дали початок всім іншим хордовим, а в покривників та сучасних безчерепних відбувається суттєва спеціалізація й спрощення органів (тупикові гілки розвитку).