ГЕНЕТИКА - Підручник - А.В. Сиволоб - 2008
РОЗДІЛ 1. Природа генетичного матеріалу
ГОМОЛОГІЧНА РЕКОМБІНАЦІЯ ДНК
Суттєвим моментом існування ДНК у живих системах є не тільки процеси відновлення та збереження інформації, яка міститься в послідовності нуклеотидів, а й різноманітні дії, спрямовані на перетасування цієї інформації з метою створення нових комбінацій генів. Найважливішою серед подібних операцій є гомологічна рекомбінація - обмін ділянками між досить довгими молекулами ДНК із гомологічними послідовностями пар основ. Такий процес відбувається в усіх організмів, котрі розмножуються статевим шляхом, між гомологічними хромосомами при мейозі (у процесі утворення статевих клітин). Також гомологічна рекомбінація можлива в парах гомологічних дочірніх молекул ДНК при реплікації в соматичних клітинах (мітотична рекомбінація) і в прокаріотів, скажімо, після кон'югації двох бактеріальних клітин і проникнення ДНК із однієї в іншу (див. розділ 5).
Необхідною умовою для здійснення гомологічної рекомбінації є гомологія послідовностей між двома молекулами ДНК по всій довжині. Загальну модель початкового етапу гомологічної рекомбінації зображено на рис. 1.26. Ініціюючою подією є дволанцюговий розріз в одній із гомологічних молекул (здійснюється спеціальними ферментами). Цей розріз за рахунок активності комплексу білків recBCD (тут і далі вказано назви білків, які забезпечують гомологічну рекомбінацію в E. coli, гомологічні білки є також і в еукаріотів) "розширюється" шляхом 5'-екзонуклеазної деградації ДНК - у результаті в місці розрізу залишаються два одноланцюгові 3'-хвости. Один із них (у комплексі з білком recA) здійснює інвазію - утворює подвійну спіраль з антипаралельним ланцюгом інтактної гомологічної молекули ДНК. Інший ланцюг цієї останньої виштовхується з дуплексу у вигляді одноланцюгової D-петлі (від displacement). Разом із 3'-хвостом D-петля здатна переміщуватись у пошуку гомології - максимальної комплементарності в межах подвійної спіралі, яка утворилася між ланцюгами двох гомологічних молекул ДНК. На наступному кроці відбувається репараційний синтез ДНК: два 3'-кінці розірваної молекули ДНК подовжуються ДНК-полімеразами з використанням як матриць двох ланцюгів інтактної молекули. До цього моменту схема на рис. 1.26 є одночасно схемою точної репарації дволанцюгового розриву в одній із двох сестринських молекул ДНК під час реплікації.
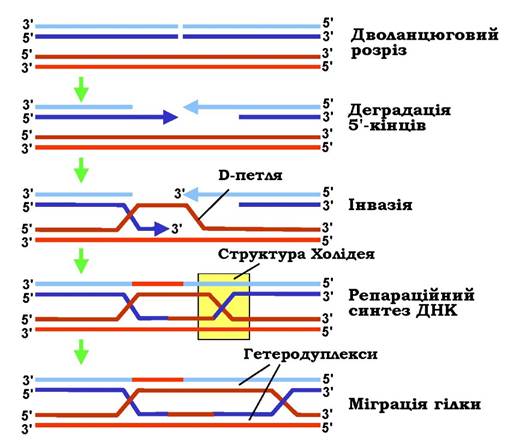
Рис. 1.26. Початкові стадії гомологічної рекомбінації
Під час рекомбінації відновлення цілісності ДНК є тільки завершенням початкового етапу. У результаті інвазії та репараційного синтезу дві молекули ДНК об'єднуються, утворюючи два перехрещення ланцюгів - дві структури Холідея (Robin Holliday). Кожна така структура може переміщуватись (так звана міграція гілки), результатом чого є подовження гетеродуплекса - подвійної спіралі між двома майже комплементарними ланцюгами двох гомологічних молекул ДНК.
Розглянемо окрему одну структуру Холідея. Як видно з рис. 1.27, її можна піддати ізомеризації, повернувши два дволанцюгові кінці на 180°. Саме у формі без перехрещення ланцюгів (праворуч на рисунку) структура Холідея фіксується завдяки її взаємодії з білком ruvA: білок зв'язується з центром хреста, утримуючи чотири одноланцюгові ділянки у приблизно планарній квадратній конфігурації. З ruvA та двома дуплексами, що виходять із хреста в протилежних напрямках, взаємодіють два гексамерні білкові комплекси ruvВ,які в АТР-залежному процесі забезпечують протягування ланцюгів через комплекс ruvА / ruvВ - міграцію гілки з одночасним подовженням гетеродуплекса.
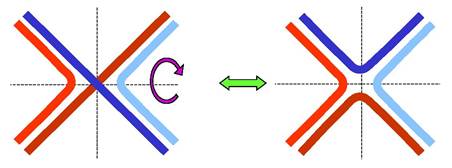
Рис. 1.27. Ізомеризація структури Холідея
Останньою подією рекомбінації є розділення (resolution) структури Холідея резольвазою - білком ruvС. Резольваза - це ендонуклеаза, дві молекули якої взаємодіють із комплексом ruvА / ruvВ і двома ланцюгами хреста, розташованими один проти одного (є два рівноймовірні варіанти такого зв'язування). Отже, резольваза робить дволанцюговий розріз через хрест двома можливими шляхами (рис. 1.28). Після наступного зшивання розривів лігазою залишаються дві дволанцюгові молекули ДНК.
На рис. 1.28 зображено ізомеризовану чотириланцюгову структуру з рис. 1.26 - дві структури Холідея з перехрещеннями перетворені на планарні (ділянки, які синтезовані шляхом репарації, не позначено) - і результат її розділення. Із чотирьох можливих комбінацій розділення двох структур Холідея показано дві. Одна з них приводить до рекомбінації (аналогічно, рекомбінантною є і пара розрізів 2 + 4): дві гомологічні молекули ДНК обмінялися ділянками, і умовна "абетка", якою позначено фрагменти ДНК, змінює регістр - великі літери замінюються на маленькі та навпаки. У масштабі цілих хромосом результатом рекомбінації є дві молекули, показані в центральній частині рис. 1.29. Пари розрізів 1 + 4 і 2 + 3 не приводять до рекомбінації. Отже, рекомбінація при розділенні структур Холідея відбувається з імовірністю 50 %.

Рис. 1.28. Схема розділення двох структур Холідея. Конфігурація чотирьох ланцюгів зверху є еквівалентною конфігурації на нижній панелі рис. 1.26. Літерами позначено ділянки ланцюгів (великі й маленькі літери відповідають гомологічним ділянкам двох молекул, літери зі штрихом і без - комплементарним ділянкам вихідних дуплексів). Цифрами 1-4 позначено можливі розрізи резольвазою. Унизу: дві пари дволанцюгових молекул після розділення структур Холідея, отримані в результаті відповідних розрізів
Незалежно від того, чи відбулася рекомбінація, усі продукти містять гетеродуплекси (середня частина всіх кінцевих молекул на рис. 1.28). Оскільки гетеродуплекси складаються з майже комплементарних ланцюгів, вони містять місметчі. Відповідно, кінцевою операцією, яка завершує процес гомологічної рекомбінації, є репарація цих місметчів описаною вище системою mutHLSU. На відміну від того, як ця система спрацьовує після реплікації, після рекомбінації ланцюг, де відбувається заміна нуклеотидів, обирається випадково, наприклад, центральний фрагмент першої молекули на рис. 1.28 перетворюється або на В/В', або на b/b'. Якщо на цій ділянці розташований ген, шляхом репарації обирається один із його алелів - В чи b. Отже, ефектом рекомбінації може бути явище конверсії гена - заміни одного алеля на інший.

Рис. 1.29. Кросинговер як результат рекомбінації двох дволанцюгових молекул ДНК
Іншим наслідком рекомбінації є обмін ділянками між двома гомологічними хромосомами - кросинговер (рис. 1.29). Під час процесу рекомбінації кросинговер між двома хромосомами може відбутися (і часто відбувається) у кількох точках (рис. 1.29). Зрозуміло, що чим більшою є відстань між двома хромосомними локусами, із тим вищою ймовірністю відбудеться обмін ділянками десь між цими локусами, і тим більшою буде кількість таких обмінів. Два локуси, що рознесені по хромосомі на велику відстань, починають поводити себе як незалежні (не зв'язані в одній хромосомі) спадкові елементи. У цьому, власне, і полягає біологічне значення гомологічної рекомбінації (детальніше ефекти кросинговеру описано в розділі 3.)
Серед інших процесів рекомбінації ДНК розрізняють сайт-специфічну рекомбінацію (вирізання / вбудовування однієї молекули ДНК з / в іншу за рахунок розпізнання специфічними білками коротких елементів послідовності ДНК, див. розділ 5), незаконну рекомбінацію (об'єднання двох молекул ДНК, які не мають ані гомології, ані специфічних елементів послідовності - основним механізмом є розглянуте вище негомологічне з'єднання кінців) і переміщення в межах геному мобільних елементів послідовності ДНК (розділ 6).