ГЕНЕТИКА - Підручник - А.В. Сиволоб - 2008
РОЗДІЛ 6. Генетика багатоклітинних еукаріотів
ЕУКАРІОТИЧНІ ГЕНОМИ
Загальні риси будови еукаріотичних геномів
Усі гени багатоклітинного організму можна розділити на дві групи: 1) гени, від яких залежать певні універсальні функції, і які є активними в усіх клітинах, - "гени домашнього господарства» (housekeeping genes); 2) гени, що специфічно активуються в клітинах певного типу, - "гени розкоші" (luxury genes). Досить загальною ознакою генів першої групи є розташування в їхніх регуляторних зонах так званих CpG-острівців - ділянок із підвищеним вмістом динуклеотидів CpG (мається на увазі послідовність нуклеотидів уздовж подвійної спіралі). Загалом вміст цих динуклетидів у еукаріотичних геномах приблизно у п'ять разів менший за очікуваний унаслідок метилювання цитозину в складі CpG-контакту: 5mC (5-метилцитозин) спонтанно перетворюється на тимін, що є одним із джерел виникнення мутацій. Метилування цитозину в регуляторних ділянках є одним із механізмів репресії генів (див. нижче). Відповідно, гени, які зберігають свою активність у більшості клітин, містять неметильовані динуклеотиди CpG, вміст яких зберігається на високому рівні.
Загальна кількість білкових генів у геномах вищих еукаріотів варіює приблизно від 20 до 30 тис. (див. табл. 1.1). Приблизний розподіл еукаріотичних білків за їхніми функціями показано на рис. 6.1.

Рис. 6.1. Приблизний розподіл білків еукаріотичного протеому за функціями
Серед еукаріотичних генів 25-50 % є унікальними (представлені в геномі єдиною копією), решта належать до родин генів, що складаються з кількох копій, як правило, не ідентичних. Відповідні (гомологічні але не ідентичні) білки становлять родину білків. Кілька родин (протеїнкінази, транскрипційні фактори певного типу, імуноглобуліни) містять сотні білків, більшість родин складається з кількох (до 30) білків. Гени такої родини часто об'єднані в геномі в кластери - знаходяться поряд у певній хромосомі (кластери генів теплового шоку, гло- бінові гени). Слід зауважити, що такий кластер не є опероном - кожен ген піддається регуляції як окрема одиниця транскрипції. Наприклад, кластер генів β-субодиниці гемоглобіну містить гомологічні гени, які активуються на певних стадіях індивідуального розвитку (рис. 6.2). Проте всі гени кластеру мають також спільну регуляторну зону, яка відповідає за загальний потенційно активний стан кластеру в клітинах, які в принципі мають здійснювати синтез гемоглобіну.
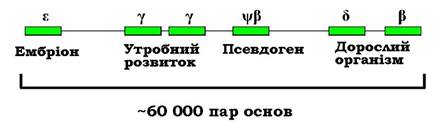
Рис. 6.2. Кластер β-глобіну в 16 хромосомі людини (кожен ген містить інтрони).
Указано стадії розвитку, на яких відповідні гени є активними
β-Глобіновий кластер містить також неактивний псевдоген (див. розділ 1). Після дезактивації гена (порушення внаслідок мутації ініціації транскрипції, сплайсингу тощо) псевдоген перестає бути об'єктом відбору, і в ньому накопичується велика кількість мутацій. Зрозуміло, що в першу чергу псевдогени з'являються саме в кластерах - коли є кілька копій гена, і пошкодження одного з них не приводить до фатальних наслідків.
Кілька генних кластерів повторюються в геномі багато разів. Серед білкових генів це стосується генів гістонів (див. розділ 1) - гени п'яти молекул гістонів завжди згруповані в кластер (кожен ген - окрема одиниця транскрипції), який повторюється до 100 разів. Іншим прикладом кластерів, що повторюються, є гени рибосомної РНК (див. розділ 2), але в цьому випадку кластер є одиницею транскрипції.
Крім генів, що повторюються, еукаріотичний геном містить значну кількість інших послідовностей, що повторюються (до 50 % геному). Основні типи таких повторів, крім уже згаданих псевдогенів, такі:
1. Тандемні повтори. До таких відносять багатократні повтори коротких послідовностей по 6-8 пар основ у теломерах і повтори так званої а-сателітної ДНК у центромерах (довжина повтору варіює від 7 пар основ у дрозофіли до 100-200 пар основ у ссавців, у людини - 171 пара основ). По всьому геному розподілені також так звані прості повтори (SSR, simple sequence repeats). Зазвичай виділяють мікросателіти - 1-15 пар основ, що повторюються від 10 до кількох тисяч разів, і мінісателіти - 15-500 пар основ, що повторюються до 100 разів. Кількість міні- та мікросателітних локусів становить десятки й сотні тисяч. Розподіл локусів за кількістю повторів є специфічною індивідуальною ознакою - на кшталт відбитків пальців.
2. Сегментні дуплікації - великі блоки довжиною 1-200 тис. пар основ, які характеризуються високим ступенем гомології. Імовірно, сегментні дуплікації є продуктом колишніх порушень хромосом. Частіше зустрічаються в перицентромерних і субтеломерних зонах.
3. Інтерсперсні (мобільні) елементи, здатні до переміщення та розмноження в межах геному, становлять основну кількість ДНК, що повторюється. Частина таких послідовностей є результатом колишньої активності мобільних елементів (таких, що втратили здатність до переміщення). Основні типи еукаріотичних мобільних елементів:
✵ ДНК-транспозони - переміщення здійснюється шляхом вирізання ділянки ДНК із наступним вбудовуванням її в інше місце - цілком аналогічно до відповідних елементів у прокаріотів. Транспозони містять один або два гени (у різних видів), що кодують транспозазу - фермент, який забезпечує транспозицію елемента, - його вирізання з донорного сайта та вбудовування в сайт-мішень. Гени транспозази можуть бути пошкодженими - тоді транспозиція даного елемента відбувається з використанням транспозази, закодованої іншим ДНК-транспозоном.
Кодуюча частина транспозона фланкована невеликими інвертованими повторами, які впізнає транспозаза, вирізаючи фрагмент ДНК. Сайт-мішень - невелика специфічна послідовність ДНК, котра теж упізнається транспозазою і теж розрізається, після чого транспозаза каталізує вбудовування транспозона до сайта-мішені (рис. 6.3). Процес транспозиції залишає дволанцюговий розріз у місці, де містився транспозон. У разі незалежної від реплікації транспозиції (нереплікативна транспозиція), цей розріз піддається репарації за механізмом негомологічного з'єднання кінців (див. розділ 1). Тобто транспозон просто "стрибає" в інше місце. Але досить часто транспозиція відбувається під час реплікації (реплікативна транспозиція) - тоді розріз репарується за механізмом гомологічної рекомбінації (див рис. 1.26): сестринська молекула ДНК використовується як матриця, і ділянка, що містила транспозон, відновлюється - транспозон і стрибає в інше місце, і залишається в донорному сайті, тобто "розмножується".

Рис. 6.3. Механізм переміщення ДНК-транспозона
✵ LTR-ретропозони - елементи послідовності, що містять довгі кінцеві повтори - Long Terminal Repeats - і кілька генів, зокрема ген зворотної транскриптази та інтегрази (аналог транспозази). Як і для наступних двох типів мобільних елементів, переміщення відбувається через проміжну молекулу РНК. Процес переміщення копії LTR-ретропозона нагадує життєвий цикл ретровірусів (див. рис. 5.9). Першим етапом є транскрипція ретропо- зона, після чого синтезована мРНК транспортується до цитоплазми, де піддається трансляції. Зворотна транскриптаза, яка є продуктом цієї трансляції, здійснює синтез ДНК: мРНК використовується як матриця, З'-кінець молекули тРНК - як прай- мер. Комплекс синтезованої ДНК-копії ретропозона з інтегразою повертається до ядра, де ця ДНК вбудовується в геном.
✵ Інтерсперсні елементи LINE (Long INterspersed Elements) містять кілька генів, включаючи ген зворотної транскриптази. Після транскрипції та наступної трансляції мРНК у цитоплазмі синтезовані білки зв'язуються з мРНК, цей комплекс повертається до ядра, де й відбувається зворотна транскрипція та вбудовування елемента в геном. Синтез мРНК при транскрипції елемента LINE, як і для більшості інших еукаріотичних мРНК, закінчується на polyA-сигналі (див. розділ 2). Цей сигнал слабкий, що дозволяє елементу вбудовуватись в інтрони звичайних генів без особливих перешкод для експресії цих генів: система процесингу часто не помічає слабкий polyA-сигнал. Елементи LINE є, відповідно, надзвичайно поширеними мобільними елементами в геномі вищих еукаріотів.
Іноді вони є не просто ділянками "егоїстичної ДНК", що автономно розмножуються в геномі, а виконують певні конкретні біологічні функції. Наприклад, у дрозофіли відсутня теломеразна система, і елементи LINE певного типу виконують функцію подовження кінців хромосом після реплікації: зворотна транскриптаза виступає як теломераза, мРНК мобільного елемента - як теломеразна матрична РНК (див. розділ 1). Цікаво, що послідовності ДНК гена теломерази та елементів LINE характеризуються високою гомологією: цілком можливо, що теломеразна система походить від мобільних елементів LINE.
✵ Інтерсперсні елементи SINE (Short INterspersed Elements) - короткі (100-400 пар основ) беззмістовні елементи, які використовують для переміщення ферменти системи LINE. До цього класу належить, зокрема, так званий Alu-повтор (назва походить від назви відповідної рестриктази, яка здатна специфічно гідролізувати цей елемент послідовності).
Мобільні елементи розподілені в геномі нерівномірно: є довгі ділянки, які на 90 % представлені мобільними елементами, і такі зони, де інтерсперсні елементи відсутні. Загалом спостерігається негативна кореляція між густиною генів і мобільних елементів. Винятком із цієї закономірності є позитивна кореляція між густиною генів та елементів типу SINE.