ГЕНЕТИКА - Підручник - А.В. Сиволоб - 2008
РОЗДІЛ 6. Генетика багатоклітинних еукаріотів
ЕУКАРІОТИЧНІ ГЕНОМИ
Генетичні ефекти активності мобільних елементів
Очевидним наслідком активності (переміщення) мобільних елементів є мутації, що спостерігаються за умови вбудовування (інсерції) мобільного елемента в кодуючу або регуляторну частину гена. Найкраще вивченими в цьому відношенні модельними об'єктами є кукурудза (саме на цьому об'єкті мобільні елементи були вперше відкриті Барбарою Мак-Клінток, див. історичну довідку наприкінці підручника) і дрозофіла.
У кукурудзи описано кілька родин мобільних елементів (Ac-Ds, Spm, Dt), які належать до класу ДНК-транспозонів. Кожна родина складається з елементів двох типів: автономних і неавтономних. Автономний елемент (одного типу на родину) здатен до переміщення за рахунок активності транспозази (див. рис. 6.3), що кодується самим автономним елементом. Неавтономні елементи, кількість і розмаїття яких є значно більшими, теж можуть переміщуватись, але тільки за умови, що вони будуть активовані автономними елементами тієї самої родини. Така активація відбувається, якщо об'єднати обидва типи елементів у геномі шляхом відповідних схрещувань. Насправді неавтономні елементи походять від автономних - це транспозони, в яких унаслідок мутацій утрачені (пошкоджені) активні гени транспозази, їхнє переміщення здійснюється за рахунок транспозази автономного елемента.
Наприклад, у родині елементів Ac-Ds роль автономного відіграє елемент Ac, а численні елементи Ds - роль неавтономних, що активуються за умови присутності в будь-якому місці геному елемента Ac (рис. 6.4). Активовані елементи Ds часто переміщуються (нереплікативним шляхом), вбудовуючись у гени, котрі мають зовнішній прояв: на рис. 6.4 показано ситуацію, коли транспозон порушує ген, який відповідає за пурпурне забарвлення зерна. У результаті виникає мозаїчність забарвлення: при наступних поділах клітини, в якій відбулась така подія, утворюється незабарвлений клон.

Рис. 6.4. Активація елемента Ds і його переміщення до гена С, який визначає пурпурне забарвлення зерна кукурудзи.
Клітинні клони, що походять від клітин, в яких відбулась така подія, втрачають забарвлення
У дрозофіли приблизно 80 % усіх спонтанних мутацій виникає внаслідок інсерції мобільних елементів. У тому числі, саме інсерція мобільного елемента copia (належить до класу LTR-ретропозонів) у локус w+Х-хромосоми приводить до мутації white, що згадувалася в розділі 3, - першої описаної мутації дрозофіли.
З активністю групи мобільних елементів дрозофіли класу ДНК-транспозонів - Р-елементів - тісно пов'язане явище так званого гібридного дисгенезу - комплексу дегенеративних аномалій, що приводять до стерильності (мутації, хромосомні аберації, порушення мейозу). Р-елементи (до 30-50 копій) містяться в геномі лабораторних ліній, які так і позначаються як лінії Р-типу, у ліній М-типу Р-елементи відсутні.
Кодуюча частина Р-елемента містить ген транспозази - 4 екзони, що піддаються сплайсингу за двома альтернативними шляхами (рис. 6.5): у статевих клітинах зріла мРНК кодує активну транспозазу; у соматичних клітинах залишається один із інтронів, що приводить до синтезу скороченого білка, який є репресором транспозицій (блокує транскрипцію на Р-елементі). Таким чином, транспозиції Р-елемента є можливими тільки у статевих клітинах.

Рис. 6.5. Схема будови мобільного Р-елемента дрозофіли (цифрами позначено екзони) і два шляхи альтернативного сплайсингу при транскрипції елемента
Звичайно, транспозиції можуть відбуватися тільки у статевих клітинах мух Р-типу. Але наявність у цитоплазмі яйцеклітини та заплідненого яйця значної кількості репресора транспозицій (материнський фактор) приводить до того, що транспозиції не відбуваються: у схрещуваннях Р-самки з будь-яким самцем народжується нормальне потомство без ознак гібридного дисгенезу. Він спостерігається тільки в нащадків від схрещування Р-самця (у якого присутні Р-елементи) і М-самки (у якої відсутні Р-елементи і, відповідно, репресор транспозицій у цитоплазмі яйця).
Цікаво, що лінії D. melanogaster, які було введено в лабораторну культуру понад 50 років тому, належать до М-типу. Мухи, знайдені в природі протягом останніх 30 років, майже завжди відносяться до Р-типу. Можливо, оскільки гібридний дисгенез запобігає схрещуванням мух різних типів, він є одним із механізмів видоутворення - сприяє ізоляції різних популяцій. Слід зауважити, що описана система є не єдиною системою дисгенезу, яка реалізується у дрозофіл.
Поряд із активністю мобільних елементів, сама наявність їх у геномі - наявність багатьох копій гомологічних ділянок ДНК - зумовлює хромосомні перебудови за механізмом гомологічної рекомбінації (див. розділ 1). Рекомбінація між мобільними елементами, які знаходяться в різних локусах двох гомологічних хромосом, є однією з причин нерівного кросинговеру, що розглядався в розділі 3, - реципрокної делеції та дуплікації хромосомних ділянок (див. рис. 3.12).
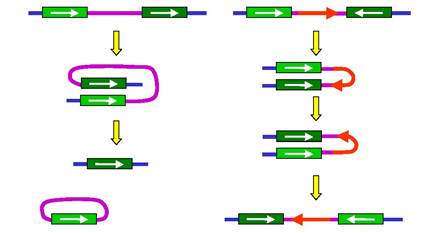
Рис. 6.6. Внутрішньохромосомний кросинговер між гомологічними мобільними елементами, орієнтованими в один бік (ліворуч,
делеція ділянки) і в різні боки (праворуч, інверсія ділянки)
Подібним чином гомологічна рекомбінація може відбуватися між мобільними елементами, що містяться в одній хромосомі. Якщо два елементи мають однакову орієнтацію своїх послідовностей, кросинговер між ними зумовлює делецію хромосомної ділянки - із хромосоми видаляється циркулярний елемент ДНК (рис. 6.6, ліворуч). Якщо орієнтація елементів є різною, відбувається інверсія ділянки хромосоми (рис. 6.6, праворуч). Як і при переміщенні мобільних елементів, результатом таких перебудов може бути втрата регуляторних ділянок, "підстановка" регуляторних елементів послідовності під інші гени, зміна напрямку транскрипції тощо.