Генетика з основами селекції - М.П.Мигун - 2008
РОЗДІЛ III. Успадкування хромосомних і нехромосомних генів
3.1.Закономірності незалежного (менделівського) успадкування
Розглядаючи цитологічні основи спадковості, ми з’ясували, як забезпечується безперервність поколінь при безстатевому і статевому розмноженні та генетичні їх відмінності (мітоз, мейоз). Аналізуючи молекулярні основи спадковості, визначили роль нуклеїнових кислот як носіїв і гарантів реалізації генетичної інформації. З’ясувати механізм запису спадкової інформації на молекулах нуклеїнових кислот та молекулярно-генетичні процеси при передачі цієї інформації і стабілізації геному.
Однак, як відбувається процес передачі спадкових ознак наступному поколінню (успадкування), цікавить людство з давніх-давен. Починаючи з кінця XVII ст. дуже багато експериментів проводилось для вивчення спадковості. Більшість учених намагалися визначити ступінь подібності батьків і нащадків одночасно за всіма ознаками. Вони виходили з того, що спадковість є неподільною властивістю організму.
Успіх Г. Менделя у цьому питанні визначається тим, що він поклав в основу генетичного аналізу принцип вивчення успадкування окремих пар ознак.
Основні особливості цього методу визначив Г. Мендель:
1. Підбір вихідних батьківських форм для схрещування; належність їх до одного виду; чіткі відмінності окремих ознак та гомозиготність із відповідних генів.
2. Використання кількісного обліку.
3. Застосування індивідуального аналізу нащадків протягом декількох поколінь.
В основі генетичного (гібридологічного) аналізу лежить метод схрещування. При генетичному аналізі для запису різних схем схрещування користуються певною символікою та правилами. Батьківські форми позначаються літерою Р (parenta - батьки). Жіноча стать - символ♀ (дзеркало Венери), чоловіча стать - ♂ (спис Марса), схрещування "X" (знак множення), нащадки - буква F (filii - діти), а покоління — F1; F2; F3;...Fn
Першою у формулі схрещування завжди вказують жіночу стать (♀А х ♂аа), тому символи статі писати не обов’язково.
Іноді результати схрещувань залежать від того, в якості материнської або батьківської використовується вихідна форма. Використовують взаємні, аналізуючі, зворотні схрещування.
Взаємні (реципрокні) - схрещування двох форм між собою в двох протилежних напрямках (АА х аа, аа х АА).
Аналізуючі - схрещування гібрида з батьківською формою, яка є гомозиготною рецесивною (дає змогу встановити структуру генотипів і гібридних форм (Аа х аа = 2Аа + 2аа), (АА х аа =Аа).
Зворотні (бекроси, насичуючі) - схрещування гібридної особини з однією з батьківських форм ( F1 Аа х АА; F2 Аа х АА; F3 Аа х АА)
Схрещування, в якому батьківські форми відрізняються по одній парі ознак, називаються моногібридним; двом парам - дигібридним; трьом і більше - полігібридним.
Вивчення явища спадковості Г. Мендель розпочав із моногібридного хрещування. Успішне застосування методу генетичного аналізу дозволило йому сформувати ряд важливих закономірностей успадкування. При моногібридному схрещуванні сортів гороху, наприклад, із забарвлення квіток, які мини червоний і білий колір, всі рослини в першому поколінні мали червоні квітки:

Якщо за материнську форму брати білоквіткові, то в першому поколінні всі рослини також були з червоними квітками:

Таким чином, незалежно від напрямку схрещування в F1 усі рослини мали червоне забарвлення квіток - одноманітні за ознакою забарвлення. Це дозволило Г.Менделю сформувати одну із основних закономірностей успадкування - правило одноманітності гібридів першого покоління.
Гібриди F1 що походять від гомозиготних батьківських особин, які відрізняються один від одного альтернативним станом певної ознаки,
є повністю одноманітними за генотиповими і фенотиповими структурами відповідної ознаки.
Ознаки, що виявляються у гібридів F1 Г. Мендель назвав домінантними та позначив великими літерами (А), а ті, що не виявляються - рецесивними, та позначив маленькими літерами (а).
Коли Г. Мендель отримав одноманітні гібриди F, за певним станом ознак, перед ним постало питання, де подівся альтернативний стан ознаки, у нашому прикладі - біле забарвлення квіток? Для вирішення цього питання він провів самозапилення гібридів F1. З’ясувалось, що серед нащадків таких гібридів у F2 поряд із рослинами, які несуть домінантні ознаки, присутні рослини з рецесивними ознаками. Підрахунки показали, що це співвідношення близько 3:1. У нашому прикладі 3 червоноквіткових і 1- білоквіткова рослина.
Закономірність у розподілі домінантних і рецесивних ознак у гібридів F2 у відношенні 3:1 називають правилом розщеплення гібридів другого покоління.
Моногібридне схрещування гомозиготних за альтернативними
формами певної ознаки особин обумовлює розщеплення нащадків F2 на дві фенотипово різноманітні за цією ознакою групи у відношенні 3:1.
Розглянемо це на нашому прикладі.
Гібриди F1 відповідно правилу одноманітності всі червоноквіткові за фенотипом, але вони утворюють яйцеклітини і спермії двох типів (А) і (а) (два типи гамет). При заплідненні, на основі рівномірного поєднання двох типів гамет, утворюється три типи зигот АА, Аа, аа.
![]()
Для спрощення аналізу результатів в F2 використовують решітку Пенета - рядки і стовпчики якої відповідають різним типам гамет, що дають батьківські форми та різним типам зигот, що утворюються при цьому:
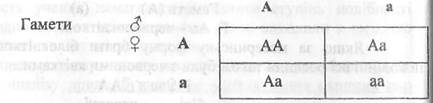
У нашому прикладі, при схрещуванні двох гібридів першого покоління, утворюються зиготи 1 АА: 2 Аа: 1 аа, тобто 3/4 червоноквіткові і 1/4 з білими квітками, що і показав Г. Мендель у співвідношенні 3:1. Указані співвідношення можливо отримувати тільки при достатньо великій кількості схрещувань.
Отже, спадкові детермінантні ознаки гібридів F1 не зникають і не зливаються, а наявні сумісно і роз’єднуються, з’являючись у черговому циклі утворення гамет.
Рослини з червоними квітками утворюються двох типів АА і Аа. АА - два однакових домінантних гени, Аа - один домінантний, а інший рецесивний, але забарвлення квіток червоне.
Організми, отримані із зигот з однаковими генами (АА) або (аа) - називаються гомозиготні, а з різними генами (Аа) - гетерозиготні.
При повному домінуванні кількість класів гібридних організмів в F2 по фенотипу і генотипу не збігається. В моногібридному схрещуванні (наш приклад) по фенотипу відокремлюється два класи: червоні і білоквіткові у співвідношенні 3:1, а по генотипу - три: АА - домінантна гомозигота, Аа - гетерозигота, аа - рецесивна гомозигота у співвідношенні 1:2:1.
Трапляються випадки, коли кількість класів за фенотипом і генотипом однакова, це відбувається при неповному домінуванні. Прослідковується у деяких випадках, наприклад, при схрещуванні червоноквіткових і білоквіткових форм нічної красуні всі гетерозиготи рожеві.

Три класи за фенотипом (червоні, рожеві, білі) та три класи за генотипом 1АА; 2Аа; 1аа. При неповному домінуванні відбувається взаємодія генів і у гібридів має місце проміжне виявлення ознаки, яке відрізняється від двох батьківських форм.
Для з’ясування суті явища одноманітності гібридів першого покоління і розщеплення ознак у гібридів F2 Г. Мендель запропонував, а У. Бетсон сформулював гіпотезу чистоти гамет. Він прийшов до думки, що розвиток будь-якої ознаки організму визначається відповідним йому спадковим фактором і передбачив наявність генів (у сучасному розумінні).
Ген - основний матеріальний елемент успадкування, ділянка молекули ДНК, що входить до складу хромосом.
Гібриди F1 і розвиваються від злиття гамет з домінантним геном (А) під червоноквіткових форм і з рецесивним геном (а) від білоквіткових - мають ген червоного і білого забарвлення квіток. Але через те, що ген червоного забарвлення домінує, то всі гібриди F1 червоноквіткові за фенотипом, однак у своєму генотипі (спадковій основі) несуть гени, які обумовлюють розвиток різних за забарвленням квіток — як червоних (А), так і білих (а).
Генотип - сукупність генетичної інформації локалізованої в соматичній клітині даного організму, його спадкова матеріальна основа.
Фенотип - сукупність ознак і властивостей, характерних для даного організму.
Злиття гамет у гібридному організмі не веде до їх змішування або знешкодження, вони залишаються такими, якими були і в батьківських організмах. У цьому і полягає суть чистоти гамет відносно однієї пари алельних генів.
Рецесивний ген (а) не зникає, не розчиняється у гібриді Аа, а зберігається в чистому стані й у подальшому проявляється у фенотипі і гомозигот (аа). Таким чином, кожна гамета, утворена Аа, утримує лише один алель - (А) або (а) у чистому стані. Те, що алельні гени у гетерозигот не імітуються і в чистому стані розходяться по гаметах, відомо як правило чистоти гамет.
Гени в гаметах, що утворили гібриди F1, залишаються такими ж окремими, якими вони були і в батьківських організмах.
Подальший розвиток генетики показав, що в гіпотезі чистоти гамет Г. Менделем було передбачено наявність генів і механізм мейозу, а також встановлено, що гени однієї пари ознак знаходяться в однакових точках гомологічних хромосом. Такі гени отримали назву алельних.
Алельні гени (алелі) - форма і стан одного і того ж гена, який знаходиться в гомологічних ділянках (локусах) гомологічних хромосом і контролює розвиток альтернативних ознак.
Поняття алелізму - одне із важливих в генетиці і явище спадковості може бути зрозумілим на основі уявлення про алельність дискретних генів.
Матеріальною основою розподілу алельних генів при утворенні гамет є процес мейозу.
Розглядаючи моногібридне схрещування, ми умовно приймали, що батьківські форми відрізняються за однією парою алельних генів. Цілковито зрозуміло, що організми різняться за багатьма генами.
Щоб одночасно аналізувати успадкування декількох пар генів, необхідно розкласти це складне явище на більш прості елементи, а після їх вивчення зрозуміти весь процес загалом. Так і вчинив Г. Мендель, прослідкувавши успадкування однієї пари ознак, він приступив до аналізу успадкування двох, трьох і більше пар ознак.
Проводячи дигібридне схрещування гороху, Г. Мендель установив третє правило закономірностей успадкування - незалежне успадкування генів (ознак).
Гени різних алельних пар і обумовлені ними ознаки передаються нащадкам незалежно один від одного і поєднуються у всіх можливих комбінаціях.
Для гібридного схрещування Г. Мендель брав гомозиготні рослини гороху, які відрізняються за двома парами ознак. Насіння було жовте гладеньке (ААВВ - домінантні ознаки) і зелене зморщене (аавв - рецесивні ознаки). Схрещування цих рослин дало нащадків Fi з фенотипом АаВb - жовті гладенькі, що підтверджує справедливість першого закону одноманітності F1.
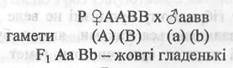
Самозапилення рослин F1 дозволяє з’ясувати кількісні співвідношення різних фенотипів і генотипів в F 2, тобто визначити характер розщеплення після схрещування Аа Вb х Аа Вb
Необхідно визначитись, які класи гамет утворюють гібриди F1, тобто батьківські форми. Це дигібридне схрещування, у якому приймають участь дві однакові дигетерозиготи (АаВb). Кожна гамета утримує два алельних гени - один із локуса А, другий із локуса В. В локусі А у гетерозиготи є два алелі А і а, а в локусі В - алелі В і b. Згідно закону випадковості їх поєднання в гаметах буде таким: (А + а) х (В + b) = АВ + Ab + аВ + ab, утворюється у дигетерозиготи чотири класи гамет:
![]()
Генотипи і фенотипи нащадків F2 знаходять математичним шляхом:
(АВ + Ab + аВ + ab) х (AB + Ab + аВ + ab) = ААВВ + ААВb + АаВВ + АаВb + ААВЬ + AAbb + АаВb + Aabb + АаВВ + АаВb + ааВВ + aaBb +AaBb + Aabb ааВЬ + aabb, або з допомогою решітки Пеннета (Табл. 3.1)
Таблиця 3.1
Розщеплення за фенотипом і генотипом при дигібридному схрещуванні
Гамети ♂ |
АВ |
Аb |
аВ |
ab |
Гамети♀ |
||||
АВ |
АА ВВ |
ААВb |
АаВВ |
АаВЬ |
жовті |
жовті |
жовті |
жовті |
|
гладенькі |
гладенькі |
гладенькі |
гладенькі |
|
Аb |
ААВb |
ААbb |
АаВb |
Aabb |
жовті |
жовті |
жовті |
жовті |
|
гладенькі |
зморщені |
гладенькі |
зморщені |
|
аВ |
АаВВ |
АаВb |
ааВВ |
ааВb |
жовті |
жовті |
зелені |
зелені |
|
гладенькі |
гладенькі |
гладенькі |
гладенькі |
|
Аb |
АаВb |
Aabb |
ааВb |
aabb |
жовті |
жовті |
зелені |
зелені |
|
гладенькі |
зморщені |
гладенькі |
зморщені |
А - ген жовтого забарвлення насіння; а - ген зеленого забарвлення насіння;
В - ген гладенької форми горошин; b - ген зморщеної форми горошин.
Аналіз даних, отриманих при розщепленні гібридів F2 в дигібридному схрещуванні, дає наступні результати:
1. За фенотипом гібриди F2 утворюють чотири класи і розподіляються в кількісному співвідношенні 9:3:3:1;
9 - жовтих гладеньких; 3 - жовтих зморщених;
З - зелених гладеньких; 1 - зелене зморщене.
2. Розподіл класів по фенотипу по кожній парі алелей проходить у співвідношенні 3:1, як і в моногібридному схрещуванні:
12-жовтих; 4-зелених (3:1)
12 - гладеньких; 4 - зморщених (3:1)
3. За генотипом дає дев’ять класів у співвідношенні: 1:2:1:2:4:2:1:2:1 1 ААВВ: 2AABb: 1AAbb: 2АаВВ: 4AaBb: 2Aabb: 1aabB: 2ааВВ: 1aabb
4. Гени кожної алельної пари (А+а) і (В+b) розподіляються на класи за генотипом, як і при моногібридному схрещуванні в співвідношенні 1:2:1
4АА:8Аа:4аа (1:2:1)
4BB:8Bb:4bb (1:2:1)
5. Відбувається новоутворення завдяки перекомбінації генів, утворюється насіння:
жовте зморщене (AAbb, Aabb); зелене гладеньке (ааВb, ааВВ)
6. Кількісне співвідношення розподілу класів за фенотипом і генотипом при схрещуванні організмів, які мають різницю за двома алельними парами, є результат добутку кількісних відношень по кожній із алсльних пар (3:1) х (3:І)=9:3:3:1 - фенотип і (1:2:1) х (1:2:1)= 1:2:1:2:4:2:1:2:1 — генотип.
Таке співвідношення фенотипових і генотипових класів за дигібридного схрещування є результатом накладання двох моногібридних розщеплень.
Суть третього закону Г.Менделя і полягає в тому, що гени (А, а, В, b) кожної із алельних пар (А+а) (В+b) у мейозі незалежно розходяться в різні гамети, а потім незалежно комбінуються в зиготах.
Цей закон підтверджується не тільки результатами дигібридного, але й тригібридного і полігібридного схрещування.
Як бачимо, кількість класів за фенотипом у моногібридному схрещуванні 2 (3:1); у дигібридному - 4 (9:3:3:1) або (22), (3:1), тригібридному - 8 (23), (З:1)3; у полігібридному — 2n, або (3+1)n, (де n - кількість алельних пар).
Кількість класів за генотипом у моногібридному схрещуванні З (1:2:1); у дигібридному 9 (1:2:1:2:42:1:2:1) або (З2), (1:2:1)2; тригібридному 27 - або 33, (1:2:1)3; у полігібридному 3n, (1:2:1)n.
Це положення відповідає для всякої кількості алелей у полігібридному схрещуванні. У полігібридному схрещуванні, знаючи кількість пар алельних генів, за якими різняться батьківські форми, можна, не залучаючи решітку Пеннета, математично розрахувати: - кількість комбінацій гамет, кількість класів за фенотипом і генотипом (табл. 3.2).
Таблиця 3.2
Розщеплення в F2 за фенотипом і генотипом для різних типів схрещувань
Типи схрещувань |
Кількість пар алельних генів (за якими різняться батьківські форми) |
Кількість гамет, що утворюється |
Кількість комбінацій гамет |
Кількість класів |
|
фенотип |
генотип |
||||
Моногібридне |
1 |
2 |
4 |
2 |
3 |
(А-а) |
А,а |
(1АА:2Аа:1аа) |
(3:1) |
(1:2:1) |
|
Дигібридне |
2 |
4 |
16 |
4 |
9 |
(A-а) (В-b) |
(22) |
(42) |
(22) |
(32) |
|
Тригібридне |
3 |
8 (23) |
64 (43) |
8 (23) |
27 (33) |
Тетрагібридне |
4 |
16 (24) |
256 (44) |
16 (24) |
81 1 (34) |
Полігібридне |
n |
2n |
4n |
2n |
3n |
Принципи спадковості і закони успадкування, які з’ясував Г. Мендель, являють собою основний зміст генетики. Їх відкриття стало умовою для поєднання всіх природничих наук у вивченні біологічних процесів спадковості.