Генетика з основами селекції - М.П.Мигун - 2008
РОЗДІЛ III. Успадкування хромосомних і нехромосомних генів
3.3. Стать і зчеплене зі статтю успадкування
Стать - це сукупність контрастуючих генеративних і пов’язаних з ними анатомічних, фізіологічних і біохімічних ознак особин одного виду.
Стать розглядається як складний детермінований комплекс ознак, який перебуває в роздільностатевих організмах у двох альтернативних станах. Стать і статеві відмінності є механізмами, які забезпечують процес комбінаційної мінливості всередині виду, а також його репродуктивну ізоляцію.
Генетика статі - це наука про генетичні закономірності визначення первинних і вторинних статевих ознак в онтогенезі, про регулювання чисельності співвідношення особин різної статі та шляхи прогнозування статі.
Ознаки, за якими відрізняються особини різних статей, поділяються на первинні і вторинні. До первинних належать ті морфологічні та фізіологічні особливості, які забезпечують утворення гамет і їх поєднання в процесі запліднення. Вторинні не беруть безпосередньої участі в утворенні гамет, спарюванні та заплідненні, але мають певне значення в статевому розмноженні.
Науково обгрунтовану відповідь, як і рішення всієї проблеми визначення і розвитку статі, було дано хромосомною теорією успадкування.
Встановлено наявність у різностатевих організмів статевих хромосом і виявлено їх роль в успадкуванні статі (X. Генкінг, 1891р.).
Однією з причин успадкування ознак із відхиленням від менделівських формул є зчеплення генів, які розташовані в статевих хромосомах (X і У) і передаються відповідно розходженню статевих хромосом - ознаки зчеплені зі статтю.
Залежно від стадії онтогенезу, на який визначається стать, розрізняють декілька типів визначення статі: прогамний тип - стать визначена ще до запліднення (в основному для видів, у яких гетерогаметна жіноча стать); сингамний - в момент запліднення і залежить від поєднання
статевих хромосом у зиготі; епігамний - після запліднення під впливом зовнішніх умов.
Існує так званий гапло-диплоїдний тип визначення статі характерний для бджіл, ос, мурашок, їздців. У них відсутні статеві хромосоми, а плоїдність визначає стать: самки - диплоїдні, а самці - гаплоїдні.
Майже весь тваринний світ поділяють на чотири типи хромосомного визначення статі (Табл. 3.3)
Таблиця 3.3.
Типи хромосомного визначення статі
Тип визначення статі |
Види організмів |
Соматичні клітини |
Гамети |
Гетеро- гаметна стать |
||
♂ |
♀ |
яйце клітини |
сперматозоїди |
|||
ХУ |
Людина тварини, дрозофіла і більшість інших видів |
XX |
ХУ |
Х і Х |
X і У |
Чоловіча |
ХУ ZW |
Птахи, метелики, рептилії |
ХУ ZW |
XX ZZ |
X і У Z i W |
Х і Х Z i ZZ |
Жіноча |
Х0 |
Коники, клопи, павуки, деякі нематоди |
XX |
Х0 |
Х і Х |
Х і 0 |
Чоловіча |
Х0 |
Міль, філоксера, попелиця |
Х0 |
XX |
Х і 0 |
XX |
Жіноча |
Якщо у соматичних клітинах однієї статі знаходяться однакові статеві хромосоми (дві X хромосоми і утворюються однотипні гамети) така стать називається - гомогаметною. В соматичних клітинах іншої статі різні статеві хромосоми (X і У) або тільки одна (X) і утворюються різні гамети, така стать називається гетерогаметною.
Кількісне співвідношення особин різної статі в нащадках близьке 1:1 і забезпечується тому, що одна стать гетерогаметна, а інша гомогаметна. Механізм забезпечення однакового співвідношення статі в нащадків, відповідає знайомому нам виду аналізуючого схрещування, коли гібрид гетерозиготний за однією алельної парою генів, схрещується з рецесивною гомозиготою Аа х аа = Аа, Аа, аа, аа (1:1) і відповідно стать ♀ХХ х ♂ХУ = 1ХХ+1ХУ.
У процесі індивідуального розвитку стать формується під впливом факторів зовнішнього та внутрішнього середовища, тому первинне (генетично визначене) співвідношення може змінюватись. Це змінне співвідношення називають вторинним. Найчастіше вторинне співвідношення зсувається в бік жіночої статі, бо у багатьох видів чоловіча стать як гетерогаметна має меншу життєздатність. У людей при народженні 100 дівчаток і 106 хлопчиків; у дитячому віці відповідно 100:103; у юнацькому 100:100; у 50років 100:85; у 85 років 100:50.
Наступним етапом за визначенням статі йде диференціація, тобто розвиток статевих відмінностей. Головним критерієм статі є формування відтворення статевої системи і фізіологічного механізму схрещування.
Встановлено, що кожна зигота є потенційно бісексуальною, тобто може розвиватися як за жіночим, так і за чоловічим типом, але є генетичні механізми, які чітко диференціюють стать за даних умов розвитку зиготи. Зачаткові гонади у ембріонів тварин є двоякої природи - індиферентні в статевому відношенні. Вони складаються із зовнішнього шару тканин кортекса (жіноча тканина) і внутрішнього - медули (чоловіча тканина). При проходженні диференціації статі і відбувається розвиток одного із задатків, але за своєю сутністю стать бісексуальна. Сам процес диференціації статі у тварин обумовлюється біологічно-активними сполуками - гормонами, які утворюються не тільки залозами внутрішньої секреції, а починають уже утворюватися в тканинах кортекса і медули. Переважання в онтогенезі гормональної секреції то однієї, то іншої статі приводить до розвитку інтерсексуальних форм.
Належність особин до тієї чи іншої статі визначається багатьма неалельними генами, що містяться не тільки в статевих хромосомах, але і в аутосомах. Тому особливості каріотипу не завжди співпадають з наявністю тих чи інших статевих ознак. В аутосомах дрозофіли знайдено рецесивний ген trans former (t), якій змінює (трансформує) стать. У гомозиготному стані цей ген жіночі зиготи (2А+2х) перетворює в фенотипових самців, але стерильних. Знайдені гени, які однодомні рослини перетворюють у дводомні. Також тож встановлено, що зміна локалізації гена може привести до порушень у прояві статевих ознак. У ссавців у У-хромосомі знаходиться домінантний ген Sex reversed (Sxr), що обумовлює розвиток чоловічої статі. Якщо ген Sxr випадково вбудовується у структуру Х-хромосоми, то із зиготи на самку (ХХ) розвивається фенотиповий самець, нездатний до сперматогенезу. Також у ссавців, включаючи людину, знайдено ген Tfm, який може викликати синдром тестикулярної фемінізації (утворення гермафродитизму).
Отже, стать контролюється цілісною системою генотипу, що передбачає взаємодію генів статевих хромосом і аутосом.
Існують докази того, що становлення статі залежить від співвідношення (дози) генів чоловічих і жіночих потенцій або різної сили її експресії. Це лягло в основу двох теорій визначення статі: балансової та фізіологічної.
Суть балансової теорії полягає в тому, що жіноча стать визначається не тільки XX хромосомами, а чоловіча ХУ. Вони визначаються співвідношенням кількості Х-хромосом до кількості наборів аутосомстатевим індексом (Х:А), при якому співвідношення Х:А рівному 1 утворюються звичайні самки; а рівному 0,5 (1Х+2А) - самці; при значенні статевого індексу більше одиниці (3Х+2А) розвиваються надсамки; менше 0,5 (1Х+3А) -надсамці; при значенні індексу меж 1-0,5 (2Х+3А) утворюються інтерсекси. Засновник цієї теорії американський генетик К. Бріджес, який встановив, що у дрозофіли розвиток ознаки статі змінюється залежно від співвідношення Х-хромосом і аутосом. Однак у більшості тварин ця закономірність не простежується.
У людини чоловіча стать незалежно від кількості Х-хромосом визначається наявністю У-хромосоми. Однак, при нерозходженні статевих хромосом у мейозі, може збільшуватись або зменшуватись кількість їх у гаметах і при заплідненні утворюються організми із захворюваннями (синдроми). Наприклад, у чоловіків синдром Клайнфельтера (А+ХХУ; А+ХХХу) (А+У - зиготи не життєздатні): безплідність, розумова відсталість, непропорційний розвиток кінцівок, високий зріст. У жінок синдром Шерешевського-Тернера (А+Х) або трисомія (А+ХХХ): безплідність, розумова відсталість, маленький зріст.
Суть фізіологічної теорії Р.Гольдшмідта полягає в тому, що стать залежить від відносної сили прояву генів чоловічих і жіночих потенцій, які завжди є у бісексуальної зиготи. Схрещуючи дві географічно віддалені раси і непарного шовкопряда - європейську і японську, які істотно відрізняються статевими потенціями (європейська раса є слабкою, а японська - сильною) з’ясувалось, що за схрещування самки європейської раси з самцями японської в F1 розвивається інтерсексуальність жіночого типу, У непарного шовкопряда Х-хромосома визначає чоловічу стать, а У-хромосома - жіночу. Жіноча тенденція У-хромосоми сильніша чоловічої Х-хромосоми, тому зигота ХУ - це зигота на самку. Однак статева потенція У-хромосоми європейського шовкопряда в сукупності з Х-хромосомою японського не дає І виразної переваги жіночої тенденції, бо чоловічі потенції Х-хромосоми японської раси дуже сильні і тому розвиваються інтерсекси.
Генетична значущість Х-хромосоми, де жіноча стать гомогаметна, І включаючи і людину, є значно більшою, ніж У-хромосоми, У Х-хромосомі дрозофіли вже картовані сотні генів, більшість із них не мають свого апеля в У-хромосомі і такі гени знаходяться в гемізіготному стані. Відповідно є гени в У-хромосомі, які не мають апеля в Х-ромосомі, їх називають голандричні. Однак, як у Х-хромосомі, так і в У-хромосомі існують гени, не пов’язані зі репродуктивною функцією. Незважаючи на те, що у більшості видів клітини жіночої стазі мають дві Х-хромосоми, а чоловічої - лише одну, але рівень експресії зчеплених з Х-хромосомою генів у самців і самок майже однаковий. Існує генетичний механізм компенсації залежної від Х-хромосоми дози генів. У плацентарних ссавців (людина, кішка, миша...) дозова компенсація здійснюється за рахунок інактивації (гетерохроматизації, лайонізації) однієї з Х-хромосом. В інтерфазних ядрах соматичних клітин самок одна із Х- хромосом концентрується біля оболонки ядра і утворює тільце Барра. Материнські й батьківські Х-хромосоми самок (при злитті гамет і утворенні жіночої статі батько завжди передає дочкам Х-хромосому відповідно і мати Х-хромосому) інактивуються в різних клітинах ембріонів за законам ймовірності. Тому самки, гетерозиготні з генів Х-хромосом, є мозаїками. Приклад кішки з черепаховим забарвленням, мають мозаїку шерсті чорних і жовтих плям. Кошенята-самці від таких кішок завжди жовті або чорні. У людей жінки, гетерозиготні за геном Х-хромосом теж є мозаїками і це проявляється при деяких захворюваннях (мозаїцизм шкіри та відсутність і зубів в деяких місцях щелеп...).
При менделівських закономірностях успадкування напрямок і схрещування, то від якої статі вноситься домінантний або рецесивний ген, не має значення для розщеплення за даними ознаками в нащадків. У такому випадку гени знаходяться в аутосомах і однаково представлені в двох статях.
Якщо ж гени знаходяться в статевих хромосомах, характер успадкування і розщеплення обумовлений поведінкою статевих хромосом в мейозі й сполученням їх при заплідненні. Гени Х-хромосоми, за деяким виключенням, не мають своїх алельних партнерів У-хромосомі. У силу цього рецесивні гени Х-хромосоми гетерогаметної статі проявляються в фенотипі (в одній дозі), бо їм не протистоять домінантні алелі у У-хромосомі.
Успадкування ознак, гени яких знаходяться в статевих хромосомах, називають успадкуванням зчепленим зі статтю. Це явище було досліджене Г. Морганом на дрозофілах і дало змогу генетично підтвердити роль хромосом у спадковості.
Оскільки статеві хромосоми гомогаметного жіночого організму передаються синам, а єдина Х-хромосома гетерогаметного чоловічого організму - дочка, то при визначенні напрямку схрещування ознаки, визначаються генами, які знаходяться в Х-хромосомі, успадковуються хрест навхрест або крис - крос успадкування. За такою схемою у людини успадковуються деякі спадкові хвороби: дальтонізм, гемофілія. Наприклад, якщо мати є носієм гену дальтонізму XXh, а батько хворий ХhУ. При готуванні таких особин ознака дальтонізму буде передаватися за такою схемою:
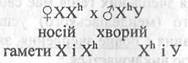
Чоловік передає дочкам завжди Х-хромосому, тому дочки з вірогідністю 25% можуть бути дальтоніками (XhXh ); 25% фенотипові моровими, але носіями гена дальтонізму (XXh); 25% синів хворими на дальтонізм (ХhУ) і 25% синів здоровими (ХУ).
Як видно із наведених прикладів у випадку успадкування зчепленого зі статтю, розщеплення ознак спостерігається вже в F1, а розщеплення в F2 може не відповідати менделівському.
Зчеплене зі статтю успадкування, не тільки не спростовує менделівських законів, але навпаки підтверджує наявність генів як матеріальних носіїв спадковості, локалізацію генів у хромосомах, обумовленість успадкування генів поведінкою хромосом у мейозі.