ТЕОРЕТИЧНІ ОСНОВИ РИБНИЦТВА - І.М. Шерман - 2011
2.ТЕОРЕТИЧНІ ОСНОВИ ФОРМУВАННЯ ТА ВИКОРИСТАННЯ ПРОДУКТИВНИХ ВЛАСТИВОСТЕЙ РИБ
2.5. Етапність розвитку риб і його використання у рибництві
Забезпечення високої рентабельності виробництва рибної продукції вимагає глибоких знань закономірностей розвитку об’єктів рибництва, а також базується на розумінні біологічних особливостей об’єктів розведення в різні періоди їх життя. Не менш важливим є також глибоке розуміння всіх без винятку технологічних процесів, особливо штучного відтворення, зокрема екологічних умов утримання плідників, стимуляції і достигання, отримання статевих продуктів, осіменіння і запліднення ікри, її інкубації, витримування вільних ембріонів, вирощування життєстійкої молоді тощо.
На сьогоднішній день найбільш розробленою теорією, яка висвітлює основні закономірності розвитку і базується на уяві єдності організму і водного середовища і на пристосувальному характері процесу розвитку, вважають теорію етапності розвитку, яка була створена В.В. Васнєцовим в 40-50-х роках минулого століття і до цього часу не втратила свого значення. Він показав, що весь розвиток риби являє собою послідовна низка етапів, кожен з яких відрізняється особливостями будови, фізіології і екології риби. Створена теорія базується на морфо-фізіолого-екологічному аналізі певного інтервалу розвитку риби майже всіх систем органів. В.В.Васнєцовим та його учнями вивчались не лише форма, а й функція тіла і плавців, шкіряних органів чуття, ротового і глоткового апарату та травної системи на послідовних стадіях розвитку. За стадію розвитку приймали кожен момент розвитку риб. Оскільки найбільш показовим критерієм був постійно зростаючий ріст риби, то вивчались личинки розміром через кожну десяту частку міліметра. Стосовно змін екології риб автори реєстрували у ембріонів відношення їх до розчинного у воді кисню, яке виражається у особливостях будови кровоносної системи і руху ембріона, у личинок і старших періодів розвитку - це перш за все зміна живлення а також зміна якості харчових об’єктів, або їх відносних розмірів. Про деякі особливості судили шляхом вивчення анатомічної будови риби. Так, наприклад, верхній рот за відповідної будови плавців слугував показником живлення біля поверхні води, нижній рот і певна форма тіла - про живлення біля дна водойм. Такий методологічний і методичний підхід і системна послідовність досліджень дозволили досить чітко спостерігати зміни у розвиткові риб.
Щоб проілюструвати, що являють собою етапи розвитку, можна навести такий приклад. Виклюнувшись із оболонки, зародки фітофільних риб, зокрема, ляща, починають плавати і підвішуватись до підводної рослинності. Вони живуть у цей час вже поза оболонки, їх екологія суттєво змінилась. Це є новим етапом у їх життєдіяльності порівняно з етапом розвитку, який відбувається в процесі розвитку ембріона всередині оболонки. Між тим ембріони ростуть і розвиваються, поступово використовуючи власний жовток. Але в ньому на цьому етапі недостатньо поживних речовин для інтенсивного розвитку і росту організму, тому виникає необхідність у додатковому живленні зовнішньою їжею. З метою забезпечення нею передличинки відриваються від рослинного субстрату, починають плавати, переслідуючи здобич у вигляді дрібних планктонних організмів, ловити її і перетравлювати. Цей новий етап в біології риб (з моменту початку живлення зовнішньою їжею називають личинкою) іменується етапом змішаного живлення, оскільки личинки поряд з власним жовтком споживають і зовнішню їжу.
Але у подальшому власний жовток личинками використаний повністю і вони переходять виключно на зовнішній (екзогенний) тип живлення, починаючи тим самим новий етап - чисто зовнішнього живлення. Автори досліджень підкреслюють, що кожній особливості біології відповідає і відповідна будова, а отже, і функція риби.
На межах етапів розвитку за переходів від одного етапу розвитку до другого, які відбуваються стрибкоподібно, з’являються зачатки нових органів і нові функції змінених органів. Так, у деяких коропових риб зачатки черевних плавців виникають при переході від шостого личинкового етапу розвитку до сьомого. При цьому ці зачатки несуть нову функцію балансирів. Протягом етапу розвитку зачатки - балансиру поступово збільшуються і укріплюються спочатку мезенхімними, а потім окостеніваючими променями, у зв’язку з тим, що риба інтенсивно росте і їй потрібні вже більш сильні балансири. У момент переходу на новий етап розвитку вони змінюють свою функцію, набуваючи здатність активного руху.
Звертає увагу й те, що групи етапів розвитку які об’єднуються загальним пристосуванням, становлять періоди розвитку. У зв’язку з цим відмічають ембріональний, личинковий, мальковий період і період статевої стиглості у риб. Характерним етапом ембріонального періоду розвитку і одночасно загальним є те, що органами дихання слугують кровоносні судини, які розташовані на жовтку, на тілі ембріона, інколи і на плавцях. При переході на личинковий період виникає зяброве дихання, але зберігаються лише залишки попередньої системи дихання. Для всіх етапів личинкового періоду загальним є те, що органами руху слугують плавцеві складки, а малькового - те, що за наявності багатьох рис будови дорослої риби відсутня ще статева стиглість, і на кінець, виникає низка особливостей, які характеризують етапи періоду статевої стиглості, у першу чергу, статева стиглість.
Слід зазначити, що В.В Васнєцов основну увагу зосередив на переходах від одного етапу до іншого етапу розвитку, на стрибкоподібності цих переходів. Між тим всі ті процеси, які відбуваються протягом всього етапу, він назвав інтервалом розвитку, протягом якого відбуваються лише повільні, поступові, майже непомітні зміни. Проте теорія етапності продовжувалась поглиблюватись і розвиватись послідовниками В.В. Васнєцова. С.Г Крижанівський встановив, що протягом кожного моменту розвитку, тобто на кожній стадії розвитку відбувається як кількісні, так і якісні зміни, і тим самим всі передумови для переходу на новий етап розвитку створюються на попередньому етапі розвитку, що підтверджує концепцію про єдність організму і середовища. Дійсно, кожен момент обміну речовин викликає і кількісні, і якісні зміни. Слід пам’ятати й про те, що ріст, який нерозривно зв’язаний з розвитком організму, становить його кількісну сторону, є результатом цього обміну.
Він також є і поступовим, і стрибкоподібним саме тому, що є результатом кожного моменту обміну речовин.
В якості підтвердження наголошуємо на тому, що споживання кожної частки власного жовтка є, навіть у незначній кількості зміна навіть форми тварини, а також зміна розміру і маси змінюють і відношення організму з водним середовищем. Таку ж картину можна уявити і по відношенню до кожної функції тварини.
Виходячи з цього, найбільший інтерес мають питання, які стосуються пояснення змін, що відбуваються в процесі розвитку організму.
Більш детальними дослідженнями виявлено, що протягом етапу поступово, послідовно накопичуються кількісні і якісні зміни будови, а отже, і функції тваринного організму і тим самим найдрібніші зміни відношень з оточуючим середовищем. Накопичення цих змін продовжується до певної межі. Але по досягненню певної міри здійснюється перехід до нової якості відношення з середовищем, перехід на новий етап розвитку, що підтверджує філософську концепцію відносно переходу кількості в якість. Щодо такого підходу цих змін досить оригінально висловився С.Г. Крижанівський, який сказав, що «... останні, зміни, які обумовлюють виникнення нової якості і перехід на наступний етап розвитку, можуть бути незначними як крапля, що переповнює чашу».
Послідовно змінюються і етапи розвитку, які складають періоди розвитку. За досягнення певної межі цих змін, здійснюється перехід до нового, більш крупного, ніж перехід від етапу до етапу розвитку, зміни якості екології тварин - перехід до нового періоду розвитку. Так, наприклад, здійснюється перехід від личинкового до малькового, від малькового періоду до періоду статевої зрілості тощо.
У розвитку загальної теорії етапності позитивну роль зіграла теорія екологічних груп риб С.Г. Крижанівського, яка дозволила усвідомити пристосувальний характер різних етапів у представників різних видів риб та представників різних екологічних груп риб число етапів розвитку, які складають періоди розвитку, виявились різними. Так, у щуки виявлено
лише три личинкових етапи розвитку, у літофітів - чотири, у фітофілів - шість. У літофільних лососевих риб личинковий період розвитку по суті може розглядатись як довготривалий етап змішаного живлення власним жовтком і зовнішньою їжею. У фітофільних коропових риб цей етап є досить коротким - протягом майже всього личинкового періоду молодь живиться зовнішньою їжею. У лососевих риб виявлено нерестовий етап розвитку, який характеризується перебудовою системи адаптації цих риб, зв’язаної з виходом на нові, сприятливі для розмноження ділянки водойми.
В природі спостерігається велике різноманіття пристосувань ембріонів риб до дихання, вони змінюються в онтогенезі і суттєво відрізняється потреби різних видів в кисні. Існують дані щодо закономірної зміни функції дихання на зародкових етапах розвитку у декількох видів риб, представників різних екологічних груп.
Встановлено також, що середні розміри риб на кожному етапі розвитку коливаються в певному діапазоні, який є специфічним для кожного виду. Однак за значних змін екологічних умов риба більш пізніх етапів розвитку може бути більш дрібною, ніж риба більш ранніх етапів. Тому недостатнім є характеристика етапу лише за розміром риб, необхідним є проведення більш глибоких і детальних досліджень з метою зібрання інформації для різносторонньої характеристики етапів розвитку.
Тривалість етапів розвитку залежить від середовища, зокрема, від наявності необхідної їжі, хімічного складу води, її температури та інших умов розвитку. Тому у різних місцях ареалу одновікова молодь певного виду може виявитись на різних етапах розвитку.
Факти розходження термінів, розмірів і морфологічних особливостей за зміни етапів розвитку у одного й того ж виду за різних умов не протирічить теорії етапності розвитку, а лише підкреслює пристосувальний характер процесу розвитку організму.
Виходячи з викладеного, доцільно розглянути періоди і етапи розвитку риб.
Теоретичні основи етапності розвитку риб, які були розроблені В.В.Васнєцовим, зводяться до того, що протягом різних періодів онтогенезу розвиток риб відбувається не лише поступово і безперервно, але і переривчасто, стрибкоподібно. При цьому відбуваються різкі зміни в будові систем органів, перебіг яких відбувається досить швидко, інколи протягом декількох годин і навіть хвилин. Ці морфологічні зміни нерозривно зв’язані із змінами біологічних особливостей риб. Між такими стрибкоподібними змінами відбувається поступовий ріст і, інколи ледве помітні кількісні зміни. Всі ці зміни відбуваються в певних межах, тому вони не змінюють основної якості, яка характеризує даний інтервал розвитку риби. Тобто якісні особливості організму і його відношення з зовнішнім середовищем зберігаються незмінними. Такі інтервали відносної стабільності у розвиткові риб між двома стрибкоподібними змінами називаються етапами.
У свою чергу кожен етап розвитку риб характеризується певними особливостями будови організму, біологічними особливостями і вимогами до умов зовнішнього середовища. За відсутності необхідних умов риби затримуються на тому чи іншому етапі. При цьому вони уповільнюють або повністю припиняють ріст і гинуть.
Спираючись на теоретичне підґрунтя, розвиток риб за теорією В.В.Васнєцова відбувається і поступово, і переривчасто (стрибкоподібно) і розпадається на низку слідуючих один за одним етапів, протягом яких відбувається ріст і поступові зміни, але не відбувається яких-небудь принципових змін в будові, фізіології або біології організму риби. Розвиток риб є послідовна зміна пристосувань їх організму до зовнішнього середовища протягом певних етапів.
Перебіг певних етапів у різних видів риб відбувається неоднаково, вони мають різну тривалість і досить часто свої специфічні особливості. Цей висновок досить важливий для розроблення біотехніки штучного розведення риб. Він вказує на необхідність вивчення біологічних і
екологічних особливостей етапів розвитку кожного виду риб, який є об’єктом штучного розведення.
Періоди і етапи розвитку риб характеризуються певними морфологічними змінами, що вимагає ретельного вивчення морфофізіологічних закономірностей раннього онтогенезу риб.
Відомо, що чисельність популяції у значній мірі визначаються особливостями раннього онтогенезу риб. Тому одним із найважливіших аспектів щодо розроблення заходів з подальшого розвитку рибного господарства важливим є встановлення закономірностей продуктивності угруповань або окремих популяцій риб та знання морфофізіологічних особливостей їх індивідуального розвитку.
Дослідженнями В.В. Васнєцова, С.Г. Крижанівського, Л.П. Рижкова встановлено, що конкретним інтервалом розвитку риб є етап, протягом якого відбувається ріст і поступові якісні зміни, які створюють необхідні передумови для переходу на нові етапи протягом досить короткого часу, але як сказано вище, стрибкоподібно. Дослідженнями було також доведено існування періодичності змін швидкості диференцировки організмів. Так, зокрема, виявлено, що посилення темпу і зростання числа диференцировок, зазвичай, спостерігається в період переходу від одного етапу розвитку до другого, тобто у так звану перехідну стадію, а їх послаблення відбувається в продовж кожного етапу. Розглядаючи онтогенез особин певного виду, а це явище загальне, розрізняють чотири періоди - ембріональний, личинковий, мальковий і статевозрілий, які характеризуються певними морфологічними, екологічними і фізіологічними особливостями. Так, ембріональний період характеризується тим, що ембріон знаходиться в оболонці яйця, личинковий період являє собою личинку з ендогенним, змішаним і суто зовнішнім живленням до появи центральних пластинок луски. Мальковий період розпочинається з утворення перших склеритів на лусці до настання статевостиглості, а статевостиглий період починається з моменту настання статевої стиглості й триває до завершення життєвого циклу. Кожен з цих періодів поділяється на ряд етапів з певними морфофізіологічними і біохімічними особливостями.
Кількість етапів у різних видів риб може бути неоднакова і залежить від їх біології. Так, наприклад, у лососей личинковий період виявлено сім етапів, у севанської форелі - вісім, що обумовлено біологічними особливостями цих видів.
Кожен з етапів у свою чергу поділяється на стадії, яка являє собою конкретну характеристику певного морфологічного і фізіологічного стану організму риб. Виходячи з цього, перехідну стадію можна розглядати як завершуючу кожен етап, яка забезпечує в результаті складних морфофізіологічних перетворень перехід на нові наступні етапи розвитку.
Спеціальними еколого - фізіологічними і біохімічними дослідженнями встановлено, що більшості видів риб в процесі онтогенезу властиві періодичні зміни фізіологічних і біохімічних процесів, зокрема, інтенсивності споживання кисню, тобто газообміну, які тісно зв’язані з морфологічними перетвореннями організму. Сутність цього явища в ранньому онтогенезі полягає у значному зростанні його величини в період перехідних стадій, і його зменшенні протягом всіх етапів розвитку. Кожному етапу розвитку властиві конкретні межі і середні рівні газообміну. Характерною особливістю також є зростання інтенсивності споживання кисню від моменту запліднення ікри й до початку живлення личинок з наступним його уповільненням за подальшого розвитку молоді риб.
Поряд з цим виявлена закономірна періодичність змін лінійного росту і накопичення маси риб, що визначається і діагностується досить чітким зростанням на початку кожного етапу розвитку з наступним уповільненням протягом етапів з мінімумом на перехідній стадії. Не дивлячись на деяке уповільнення швидкості росту організмів протягом етапів, кожному етапу розвитку властиві конкретні межі змін і середні величини швидкості, які обумовлені морфофізіологічними особливостями організмів протягом даного періоду розвитку, видоспецифічними особливостями риб.
Загальною закономірністю є те, що кожний якісно новий етап розвитку починається із значного зростання швидкості росту і потім також демонструє уповільнення. У той же час на початку етапу інтенсивність газообміну відносно стабілізується на рівні, характерному для відповідного етапу. Його величина в 1,5-3,0 рази нижче, ніж в період перехідної стадії. Протягом етапу темп диференцировок незначний. Під кінець етапу як швидкість росту, так і інтенсивність газообміну, досягає максимальних величин, але темп диференцировок може дещо зростати. У цілому організм у цей час підготовлений до переходу на новий етап розвитку. В результаті морфофізіологічних і біохімічних перетворень під час перехідної стадії відбуваються якісні зміни в організмі, який переходить на якісно новий етап. Перехідна стадія розпочинається з різкого зростання інтенсивності споживання кисню, який забезпечує необхідні енергетичні можливості для морфологічних та інших перетворень. За досягнення максимального рівня дихання інтенсивно зростає темп диференцировки. У цей час лінійний і ваговий ріст організму досягає свого мінімуму. Всі енергетичні можливості організму спрямовані на швидке забезпечення перехідної стадії, протягом якої зареєстровано недостатньо стійка його взаємодія з оточуючим середовищем. Під кінець перехідної стадії інтенсивність газообміну і темп диференціації уповільнюються і до початку етапу відносно стабілізуються, а швидкість росту організму зростає. Зміни, які відбуваються протягом етапу розвитку і в період перехідної стадії, представлені на рисунку 2.8.
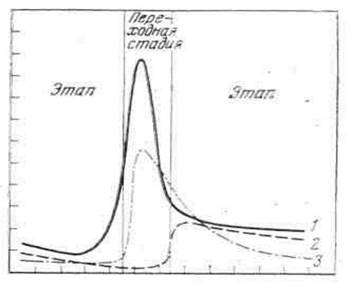
Рис. 2.8. Зміна швидкості росту і інтенсивності газообміну за переходу на нові етапи розвитку (за Рижков Л.П., 1984)
1 - інтенсивність газообміну; 2 - швидкість росту; 3 - темп диференціації.
Поглиблене вивчення періодичності перебігу процесів диференціації організму, швидкості росту і інтенсивності газообміну має не лише теоретичний інтерес, але й практичне значення для вивчення продуктивності біологічних систем і раціонального використання їх продукції за умов організації досліджень на якісно новому енергетичному рівні.
У процесі розвитку риб до досягнення статевої стиглості встановлено зростання абсолютних показників пластичного і функціонального обміну. Це може бути обумовлено загальним зростанням маси тіла риб, яке вимагає в процесі розвитку підвищення потреб енергетичних ресурсів як для побудови тіла організму, так і для здійснення основних процесів життєдіяльності. Загальною закономірністю для більшості прісноводних риб є суттєве зростання швидкості функціонального обміну порівняно з пластичним. Зміни величин функціонального і пластичного обміну тісно взаємозв’язані як між собою так і з інтенсивністю процесів морфогенезу, оскільки відомо, що найбільш інтенсивно морфологічні перетворення відбуваються в період переходу від одного етапу розвитку до другого - під час перехідної стадії.
Встановлено, що при переході на новий етап розвитку відбувається різке і, як правило, значне зростання використання трансформованої
енергії їжі на функціональний обмін, який забезпечує необхідною енергією морфологічні перетворення, свідченням чого є зростання темпу диференцировок і морфологічних змін.
Біологічна доцільність збільшення енергетичних витрат на функціональний обмін полягає в необхідності якнайшвидшого забезпечення переходу організму в якісно новий стан і тим самим ліквідації деякої нерівноваги, яка виникла між організмом і середовищем в кінці попереднього етапу. Після переходу на новий етап розвитку інтенсивність функціонального обміну знижується, а величина пластичного обміну зростає. Тобто в цей період відбувається перерозподіл загальної енергії їжі на різні процеси обміну речовин, які мають в кожний конкретний момент найбільш важливе значення для розвитку і росту організму. У деяких випадках відбувається одночасне збільшення абсолютних показників функціонального і пластичного обміну на початку нового етапу розвитку, що може бути пов’язано з значними ускладненнями структури організму і досить суттєвими змінами їх біології. Так, на етапі розвитку початку змішаного живлення зростання функціонального обміну може бути пов’язано із збільшенням витрат енергії організму на пошук їжі, її захоплення, перетравлювання тощо.
Протягом кожного окремого етапу розвитку пластичний обмін, зазвичай, переважає над функціональним, величина якого до кінця етапу знижується. Ця закономірність посилюється можливим вичерпуванням можливостей як морфологічних, так і функціональних конкретного етапу розвитку організму. Тому виникає необхідність нових морфофізіологічних і біохімічних перетворень для переходу організму на наступний етап розвитку.
Поряд з розглядом загальних закономірностей раннього онтогенезу риб в умовах господарської діяльності виключне значення має ембріональний і личинковий періоди розвитку, що орієнтує на доцільність окремого розгляду.
Запліднена ікринка - це вже ембріон, який розвивається, проходячи низку певних етапів розвитку. На етапі запліднення після проникнення сперматозоїда в ікринку спостерігаються складні перетворення і переміщення її вмістимого. Через 12-15 хв. після осіменіння ікринки деяких видів риб (коропових, осетрових) стають клейкими, які легко приклеюються до субстрату. Оболонки ікринок набрякають у воді, стають прозорими і міцними. Ікринки значно збільшуються у розмірах. Питома маса ікринок зменшується. Перивітелліновий простір заповнюється рідиною. Поверхневий шар цитоплазми зміщується у напрямку до протилежного боку анімального полюса. Цей етап закінчується злиттям (поєднанням) жіночого і чоловічого пронуклеусів. Перехід до наступного етапу розвитку - етапу дробління - починається з моменту появи на ікринках першої борозни, яка утворює два бластомери. На цьому етапі кожна ікринка послідовно ділиться на все більшу кількість бластомерів, а в кінці його досягає стадії бластули. Далі зародок вступає на новий етап розвитку - етап гаструляції, за якого починають формуватися тіло зародка і його жовтковий мішок. На наступному етапі розвитку зародка відбувається нейруляція. У зародка утворюється нервова трубка, яка являє собою зачаток нервової системи.
У подальшому у ембріона змінюється форма тіла, обособлюється хвостовий відділ, формуються відділи головного мозку, закладаються органи чуття і залози викльовування. У ембріона починають формуватися травна і видільна системи, утворюються серце і кровоносні судини, закладаються зяброві кишені, з’являється зачаток печінки, утворюються м’язові волокна, клітини крові і починається вакуолізація клітин хорди. Тіло ембріона розчленовується на окремі сегменти і на ньому з’являється плавцева складка.
Ембріон, який пройшов низку етапів розвитку, вже має функціонуюче серце і кровообіг, а також нервову систему і відповідає на зовнішні подразнення. У цей час міцність оболонок ікринок послаблюється під дією ферментів вилуплення і ембріони звільняються від них.
Ембріональний період закінчується на етапі, за якого передличинки, які вилупились, завершують свій розвиток і стають личинками.
З першого етапу свого розвитку ембріон вступає в тісний контакт з оточуючим середовищем. Значний вплив на розвиток ембріона (ембріогенез) має температура води, її газовий режим, рН, солоність, осмотичний тиск та освітленість. У кожного виду риб ембріогенез відбувається у певних температурних межах. При цьому розрізняють температурний оптимум, температурний поріг і температурний максимум. Температура, за якої ембріогенез відбувається нормально, називається температурним оптимумом. Підвищення температури прискорює процеси ембріогенезу, а зниження - уповільнює цей процес.
Температура, нижче якої ембріогенез не відбувається, називається температурним порогом. Однак порогова температура не приводить до загибелі зародків за впливу протягом обмеженого впливу часу. За відновлення оптимальної температури ембріогенез відновлюється і відбувається нормально. Температура, вище якої ембріогенез припиняється і ембріон гине, називається температурним максимумом. Знання температурних меж, в яких відбувається нормальний ембріогенез, важливо не лише для більш точного прогнозування майбутньої рибопродуктивності, але й для правильного ведення процесу інкубації ікри за штучного риборозведення.
Не менш важливу роль на процес формування ембріона має і газовий режим. За відсутності кисню у воді зародок гине від задухи. Знижений вміст кисню у крові уповільнює розвиток ембріона. Якщо ж недостатність кисню проявляється за підвищеної температури (порівняно з оптимумом), то розвиток ембріона відбувається не нормально, що приводить до утворення потворних особин, а у подальшому - до їх загибелі. Подібне явище спостерігається і за високих концентрацій СО2, які можуть бути за інкубації великої кількості ікри і у невеликому об’ємі непротічної води. У цьому випадку СО2 зсовує активну реакцію середовища в кислий бік і порушує газовий обмін в клітинах зародка.
Відношення ембріонів до температури, газового режиму, солоності, світла, механічних впливів на окремих стадіях етапів розвитку неоднакове. Одні стадії більш чутливі до різких змін абіотичних умов середовища, що приводить до збільшення кількості потворних ембріонів і їх загибелі, інші - менш чутливі. Підвищена чутливість до різких змін водного середовища проявляється на стадіях перших етапів (запліднення-гаструляція) і на етапі перед викльовом.
Отже, інкубацію ікри можна проводити лише в таких умовах, за яких фактори зовнішнього середовища забезпечують нормальний перебіг ембріогенезу. За цих умов зародок добре росте і розвивається, використовуючи поживні речовини жовтка.
Ембріон, що виклюнувся, або так звана передличинка, перший час веде пасивний спосіб життя. Вона живиться за рахунок поживних речовин, які містяться в жовтковому мішку, який являє собою тимчасовий орган. Жовтковий мішок є також провізорним органом дихання передличинки. По мірі росту передличинки жовтковий мішок поступово зменшується. Невдовзі до кінцевої його резорбції закінчується ембріональний і починається личинковий період розвитку. Передличинка стає личинкою, котра переходить на змішаний тип живлення. На цьому етапі розвитку личинка використовує поживні речовини, які містяться у жовтковому мішку, і споживають їжу із зовнішнього середовища.
Жовтковий мішок у личинки невдовзі повністю резорбується і вона повністю переходить на зовнішнє живлення. Личинка росте, розвивається і через деякий час перетворюється у малька, який за зовнішнім виглядом нічим не відрізняється від дорослої риби.
Тривалість личинкового періоду розвитку, як і ембріонального, у різних видів риб неоднакова - від декількох діб до місяця. У кожного виду риб вона зростає або зменшується в залежності від температури води та інших чинників абіотичних і біотичних умов середовища.
Знаючи етапи розвитку риб, можна вивчити шляхи і функціональні механізми перетворення популяцій прохідних, напівпровідних і прісноводних риб у сучасних умовах комплексного використання водних біоресурсів на основі виявлення максимальних можливостей пристосованості цих риб до змін, які відбуваються у водному середовищі.
Запаси жовткового мішку витрачаються передличинками з різною швидкістю, що значною мірою залежить від температури оточуючого середовища. В цьому зв’язку доцільно мати відповідне уявлення відносно впливу температури на ріст і використання жовтка в ранньому онтогенезі риб.
Зазвичай, в іхтіології ріст ембріона і динаміку використання жовтка в ембріональний період визначають традиційними методами, зокрема, шляхом вимірювання довжини, маси ембріонів, маси жовтка. У деяких випадках здійснюють розрахунок об’єму ембріона і жовтка або площ їх проекцій.
Між тим існує визначення Ф.Р.Хейса про те, що ріст ембріона - це, перш за все, утворення і накопичення в його тканинах білка, яке є підґрунтям зростання його маси і лінійних параметрів.
Дослідженнями Г.Г. Новікова та Н.Д. Куфтіною встановлено, що ріст білкової маси ембріонів тріски, пінагора, благородного лосося на перших етапах ембріогенезу за різних температурних умов мінімальний, що пояснюється низьким рівнем білкового синтезу в бластодермі. Цю думку поділяють Нейфах А.А., Тимофеева М.Я. (1977) базуючись на відповідних дослідженнях.
Деяке зростання білкової маси починається з етапу гаструляції (у тріски і пінагора), а у лосося - з етапу органогенезу. У подальшому швидкість білкового росту зростає, у тому числі і з підвищенням температури води. Спеціальними дослідженнями встановлено, що чим вище температура розвитку, тим більша швидкість білкового росту протягом доби. Але в момент виходу ембріона із оболонки величина його білкової маси з підвищенням температури знижується.
Роботами науковців показано, що з підвищенням температури тривалість розвитку ембріона скорочується у більшій мірі, ніж зростає швидкість росту білка. Саме цей взаємозв’язок може бути однією з причин зменшення кінцевих розмірів зародків в момент їх виходу із оболонки за більш високих температур. Це свідчить про те, що нормальний розвиток ембріона відбувається за оптимальних температурних умов, а порушення динаміки розвитку може розглядатись як вихід температури за межі оптимальних величин.
Дослідження показали, що, процес розвитку ембріона і його життєдіяльність здійснюється виключно за рахунок запасних речовин жовтка, основним компонентом якого є білкові речовини (Нейфах А.А., Тимофеева М.Я.) (1977).
На відміну криволінійного характеру росту тіла зародка, швидкість якого по мірі росту змінюється, процес резорбції білкових речовин у жовтку відбувається з постійною швидкістю і має вигляд прямолінійної залежності. Враховуючи ці закономірності, автори досліджень дійшли висновку про те, що процеси регуляції швидкості росту білкової маси тіла зародків і резорбції запасних білків жовтка можливо мають відносну незалежність.
Слід зазначити, що швидкість резорбції запасних речовин, як і швидкість росту ембріону, з підвищенням температури зростає. При цьому також виявлено, що характер резорбції білкових речовин жовтка погоджується з активністю протеолітичних ферментів, які здійснюють розщеплення білкових молекул жовтка. У нормі активність кислих протеаз залишається незмінною протягом розвитку ембріону, проте з підвищенням температури води вона зростає відповідно швидкість резорбції жовтка.
Не дивлячись на те, що з підвищенням температури розвитку швидкість резорбції білка в жовтку зростає, загальна кількість білка, резорбованого за період розвитку ембріона під оболонкою, навпаки, зменшується. Тобто величина залишкового білка в жовтку на вихідних стадіях розвитку знаходиться в прямій залежності від температури.
Наслідком непропорційних змін швидкостей резорбції білка в жовтку, з одного боку, і тривалості розвитку ембріону, з другого за підвищених температур є те, що зародки, що розвиваються за цих температур, виходять з оболонки менших розмірів і з більшим запасом білка в жовтку. Це може свідчити про порушення процесів ембріонального розвитку риб за дії підвищених температур.
Заслуговують на увагу дослідження, пов’язані з вивченням динаміки вмісту білка в жовтку в процесі розвитку ембріона, яка встановлюється шляхом вивчення зміни білка в цілій ікринці. Ця динаміка визначається, з одного боку, масштабами резорбції білка в жовтку, а з іншого - ростом білкової маси зародка. Дослідженнями И.Е. Мороза та В.П. Лукіна (1973) показано, що на початку розвитку, коли ріст ембріона мінімальний, вміст білка визначається лише резорбцією білка в жовтку. З початком інтенсивного білкового росту вміст білка в ікринці зменшується і в кінці ембріонального розвитку стабілізується.
Наукові розробки у складі авторів Г.Г. Новікова та Н.Д. Куфтіної (1988) свідчать, що у цілому використання білка жовтка в процесах розвитку ембріона коливається в межах 10-40 % і залежить від виду риб і температури води. В процесі розвитку ембріона пінагора, тріски і лосося під оболонкою резорбується біля 30-40 % білка, але на ріст безпосередньо ембріона використовується 40-80 % резорбованого білка у тріски і пінагора, а у лососей - лише від 20 до 30 %. Це свідчить про те, що масштаби білкового росту ембріонів на одиницю вихідної білкової маси, ікри в процесі розвитку під оболонкою у тріски і пінагора в 2-3 рази вище ніж у благородного лосося.
Білок, який залишається, у значній мірі використовується організмом після виходу ембріона з оболонки. Тому у риб з дрібною ікрою і невеликою кількістю запасних речовин до початку екзогенного живлення використовуються практично всі запасні речовини жовткового мішка. Саме тому передличинки цих видів риб не можуть голодувати протягом тривалого періоду на відміну від передличинок лососевих риб, які мають у жовтковому мішку більшу кількість резервних речовин у яких після викльовування реалізується біля 70-80 % запасного білка.
Виходячи з цього великий вміст в ікрі запасних резервних речовин, спрямований на забезпечення розвитку організму поза оболонки ендогенними ресурсами до переходу на екзогенний тип живлення.
В життєвому циклі риб та в процесах розведення значні втрати відбуваються при переході від ембріону до личинки, що має наукове підґрунтя і підтверджується практичним досвідом. Об’єктивна реальність орієнтована на доцільності вивчення закономірностей переходу від ембріонального стану до личинкового. Торкаючись цього питання доцільно нагадати, що у онтогенезі риб розрізняють два ранніх періоди - ембріональний і личинковий, які складаються з етапів, що поступово змінюють один одного. Кожен з цих етапів характеризується цілим комплексом морфологічних, фізіологічних, біологічних ознак, які виникають в результаті реалізації спадковості за конкретних екологічних умов. Кожному етапу розвитку властивий певний комплекс органів і їх функцій, які змінюються в залежності від впливу чинників оточуючого середовища. Проте ці зміни мають свою межу, яка є наслідком реалізації адаптивних можливостей організму, вироблених в процесі еволюції виду.
Доведено, що процес ембріонального розвитку організму у кожному конкретному випадку обумовлюється дією багатьох чинників, що в кінцевому рахунку приводять до різноякісності нащадків, які мають більш широкий діапазон адаптацій, що забезпечують збереження оптимальної чисельності популяції. До числа чинників, які прямо або опосередковано впливають на ембріогенез, відносяться: вік і фізіологічний стан плідників; місце розташування ікри в яєчнику і ступінь її стиглості; термічний режим в період овуляції; статева активність сперміїв; термічний режим в період ембріонального розвитку, який впливає на диференцировку ембріона і його метаболізм, а також мінералізація води, її активна реакція, пігментація ікри тощо.
Ембріональний період закінчується виходом ембріона із яйця, після чого починається личинковий період, перший етап якого - це етап ендогенного живлення за рахунок резервних речовин, зосереджених у жовтковому мішку.
Момент виходу ембріона з яйцеклітини на перший погляд створює враження короткочасного стрибка. Однак у дійсності викльовування ембріона є досить тривалим процесом, який супроводжується накопиченням суттєвих морфо-фізіологічних змін. Цей процес складається із підготовки ембріону до викльовування, самого моменту викльовування і тих змін, які забезпечують найголовніші процеси життєдіяльності організму, який виявляється поза яйцевої оболонки. Кожен з цих моментів у кожному конкретному випадку може бути зсунений у будь-який бік дією температури, газового режиму, освітленості, механічного впливу.
Одним з критичних періодів ембріонального розвитку є підготовка ембріона до викльовування. У цей час ембріон є найбільш чутливим до зовнішніх впливів. Підвищена чутливість у цей період пояснюється досягненням певного для даного виду рівня розвитку комплексу органів, інтенсифікацією ембріональної моторики, посиленням гальмівного впливу яйцевої оболонки на газообмін ембріона, накопиченням в залозах вилуплення ферменту, який розчиняє яйцеклітинну оболонку. Між тим, шляхом зміни температурного режиму, концентрації розчиненого у воді кисню чи вуглекислоти та зміною інших чинників можна порушити погоджений хід ембріогенезу, викликати затримку викльовування ембріону або, навпаки, стимулювати його передчасний вихід, що є не бажаним.
Одним з найбільш суттєвих чинників, які впливають на викльовування, є температура води, її активна реакція, вміст розчинених у воді газів, механічні впливи тощо. Досить важливим висновком деяких досліджень виявилось те, що передчасне викльовування у риб призводить до загибелі ембріонів - личинок, коли вони досить рано покидають яйцеву оболонку.
За іншої ситуації, коли викльовування настає тоді, коли вже склався комплекс життєво важливих органів (завершилася сегментація тіла,
головний і хвостовий кінці тіла відділились від жовтка, з’явилась пульсуюча серцева трубка, яка проштовхує безколірну плазму, розвинулась нервово-м’язева моторика) за таких умов ембріони, які виклюнулись, виявляються життєздатними.
Між тим, в природних умовах вилуплення затримується з різних причин, особливо за зниження температури, опосередкована дія якої змінює газообмін ембріону, знижує напруженість ембріональних рухів і активність ферменту викльовування.
Слід звернути увагу на те, що пристосування, які забезпечують виживаність ембріонів поза яйцевої оболонки, повинні бути більш широкими у видів, які розвиваються за умов різких коливань середовища, особливо на мілководдях у ранньовесняний час. Існує інформація про значну пластичність пізніх ембріонів і ранніх личинок, яка виникає як адаптація. Ця пластичність полягає у тому, що у передличинок, а в подальшому і у личинок, які виклюнулись передчасно, прискорено розвиваються життєво важливі органи, особливо прогресує у розвиткові кровоносна система, яка забезпечує у цей час інтенсивний обмін речовин ембріона, який виявився в нових умовах. Доведено також те, що чим слабкіше розвинена личинка, тим темпи її розвитку вищі. Прискорений розвиток органів відбувається до моменту утворення комплексу органів, властивих ембріона при викльовуванні за оптимальних умов розвитку.
Тривалість першого етапу розвитку - стану ендогенного живлення - буде залежати від ступеню сформованості ембріона в момент викльовування. У одних період жовткового живлення буде більш тривалим, у інших - коротким. Крім того, личинки, що виклюнулись, як більш активні, будуть споживати жовток швидше, ніж однойменні ембріони, які залишилися під яйцевою оболонкою.
Отже, викльовування можна розглядати як стрибок, тривалість якого буде залежати як від видових адаптацій, так і від конкретних умов розвитку.
Біологічні і екологічні особливості розвитку риб на різних етапах суттєві складові рибогосподарської науки, які мають теоретичне і практичне значення. Знання етапів розвитку сьогодні дозволили вченим рекомендувати рибним підприємствам використовувати молодь анадромних риб до покатного стану, перехід до якого супроводжується складними змінами, які готують організм до існування в нових умовах - в умовах життя у морі, зокрема, лососевих. Для вирішення практичних завдань, які стоять перед штучним риборозведенням, велике значення має вивчення чинників зовнішнього середовища, які впливають на організм риб на різних етапах розвитку. Все це сприяє розробленню найбільш ефективних прийомів розведення риб.
Вчення про етапність розвитку риб виявилося найважливішим теоретичним підгрунттям для розробки біотехніки вирощування молоді промислово цінних видів риб і сприяє підвищенню ефективності штучного риборозведення, що орієнтує на набуття певної інформації відносно особливостей нерестового етапу і його специфіки у розвитку риб.
Знання біології розмноження риб, їх фізіологічних особливостей і екологічних вимог в період ікрометання необхідні з метою уточнення рибоохоронних заходів, підвищення ефективності робіт з меліорації нерестовищ, для розробки екологічного способу стимулювання визрівання плідників, підвищення ефективності нересту різних видів риб в нерестово - вирощувальних господарствах, в селекції плідників тощо.
Як відомо з викладеного вище, теорія етапності, створена В.В. Васнєцовим, базується на уяві про нерозривний у протирічній єдності організму і середовища і розглядає розвиток організму як пристосувальний процес. Згідно цієї теорії процес розвитку є і безперервним, еволюційним, і у той же час перервним і стрибкоподібним, а весь розвиток організму відбувається поетапно. Протягом певного відрізку часу, який зветься етапом, біологія організму, який розвивається, суттєво не змінюється. В.В. Васнєцов писав, що: «Кожний етап відрізняється від інших специфічною для нього будовою, фізіологією і біологією, причому і будова, і фізіологія пристосовано до даної біології і до даних специфічних умов». Тобто для кожного етапу характерна певна система адаптацій. Зміни протягом етапу, які носять як кількісний, так і якісний характер, не переходять певної межі, не приводять до зміни системи адаптацій, однак готують до цього організм, що розвивається. При переході на новий етап розвитку, який здійснюється стрибкоподібно, суттєво змінюється вся біологія організму, а біологічно подібні етапи об’єднуються в періоди розвитку.
Досить цікавим в цьому аспекті є питання, пов’язане з інтервалом часу, протягом якого здійснюється нерест риби. Відомо, що багато видів риб для ікрометання обирають специфічні місця, мігрують до певних ділянок водойм. Літофільні риби йдуть виметувати ікру на ділянки з підводною рослинністю, хоча місця нагулу їх знаходяться в інших ділянках. Риби, які виметують пелагічну ікру в прісній воді, для ікрометання виходять на течію. Є риби, які нагулюються у прибережній частині моря, а для розмноження йдуть в пелагіаль. Тихоокеанський оселедець і деякі інші види, які є пелагічними, для розмноження підходять до берегів. Суттєво відрізняється нерестова обстановка у прохідних осетрових, лососевих та інших видів риб. Виходу риби на екологічне своєрідне нерестовище передує функціональна і зв’язана з нею морфофізіологічна перебудова організму.
Однак, різкі відмінності місць розмноження і нагулу властиві далеко не всім рибам і тому переднерестові зміни виражені слабо або зовнішньо можуть бути непомітними.
Є й такі види риб, які в період розмноження не змінюють ділянки водойми, однак у них все ж - таки відбуваються суттєві морфологічні зміни, обумовлені специфікою біології розмноження. При цьому змінюються морфологічно й функціонально різні органи і тканини, а також з’являються нові органи і функції. У яйцеживородячих, живородячих і виношуючих риб перетворюється кишечник, статеві та інші органи, які забезпечують постачання ембріонів, що розвиваються, не лише киснем, але й поживними речовинами. Завдяки різним новоутворенням, які розвиваються в переднерестовий період, і зміні функцій різних органів риба вступає з абіотичним і біотичним середовищем, зокрема з особинами протилежної статі і своїми нащадками, в якісно нові відношення і суттєво змінюється її біологія.
Суттєвий вплив на організм риб в період нересту має її відношення до їжі. Багато видів риб в період розмноження припиняють живитись. Це риби, які йдуть розмножуватися в місця, які не забезпечують плідників їжею, а характерними представниками таких є лососеві, осетрові. Окремі види, які виношують ікру в роті або в кишківнику, також не живляться в цей період. У інших видів риб змінюється склад харчових організмів у зв’язку з різноманіттям фауни місць розмноження і нагулу. Змінюється також ритм живлення у зв’язку із зміною всієї біології риб, що розмножуються. Припинення живлення, зміна їжі і ритму живлення призводить до зміни характеру обміну. Відомо, що обмін речовин в період розмноження змінюється також внаслідок зміни діяльності залоз внутрішньої секреції. Накопичені протягом нагулу резервні речовини спрямовуються на формування статевих продуктів, а у живородячих риб і на формування самих ембріонів; спостерігається генеративний обмін.
При виділенні етапів розвитку великого значення надається зміні способу добування їжі, характеру живлення і обміну речовин.
Переднерестовий, і, особливо нерестовий період, характеризується зміною характеру зграй і формування особливих нерестових угруповань, розвитком в шлюбний період складних поведінкових реакцій, що свідчить про вступ риб, що нерестять, в специфічні відносини, які властиві саме цьому періоду.
Отже, розмноження риб не може здійснюватись в межах попередніх адаптацій. Одним із способів вирішення цього протиріччя є перебудова організму. Зміни, які відбуваються в організмі риби за її переходу в нерестовий стан, означають зміну пристосувань, які необхідні для забезпечення процесів розмноження. Ця перебудова готується поступово і завершується стрибкоподібно перед початком ікрометання.
Стрибкоподібний характер зміни біології є одним з важливих ознак переходу організму на новий етап розвитку.
Оскільки риби в період нересту відрізняються фізіологічним станом, будовою, поведінкою, входять у специфічні відношення з абіотичним і біотичним середовищем і виконують особливу функцію, цей відрізок онтогенезу виділяється в якісно новий, нерестовий етап розвитку.
Поряд з викладеним у загальному плані доцільним є більш детальний аналіз цього етапу у моноциклічних і поліциклічних риб, враховуючи наявних особливостей відтворення.
Моноциклічні риби, яскравими представниками яких є тихоокеанські лососі, вугрі, деякі бички, після нересту гинуть або іншими словами відтворення відбувається один раз протягом життя.
У риб, які розмножуються протягом життя багаторазово, поліциклічних, онтогенез включає декілька циклів розвитку гонад.
Вчені застерігають, що повторність нересту не можна плутати з сезонною циклічністю, яка відображує особливості середовища всіх організмів, що розвиваються. Відомо, що кожна популяція розмножується в певний сезон року. Однак не кожна особина розмножується з настанням даного сезону. До нересту приступають лише фізіологічно підготовлені особини, що свідчить про відносність зв’язку нерестового етапу з сезонною циклічністю.
Між тим важливим є те, що нерест у моно- і поліциклічних риб завершується принципово однаково-закінченням циклу розвитку гонад і переходом риби в новий біологічний стан: у перших це буде смерть, а у других - перехід до нагулу. Тобто цей факт та інші свідчать про відсутність принципових відмінностей між моно-і полі циклічними рибами.
Однак у деяких видів чи особин риб існує проблема зміщення термінів статевого вистигання під впливом оточуючого середовища. Але досить важливим є встановлення цих чинників водного середовища, які визначають цей момент, що пов’язано з розробленням методів
прискорення або гальмування процесів статевого визрівання, тобто з управлінням процесами розмноження риб.
Між тим вже зараз відомі деякі з чинників, які сприяють вистиганню риб. Завершення вистигання і перехід плідників у текучий стан нормально відбувається за впливу на організм факторів водного середовища, які визначають екологічну специфіку місць ікрометання виду. Тобто підготовка і перехід риби до нерестового стану, як і увесь процес розвитку, - процес пристосувальний.
Відомо, що для переходу риб в текучий стан, в умовах штучного відтворення для певних видів риб, застосовують фізіологічно активні речовини. Однак цей метод фізіологічного або гормонального стимулювання вистигання риб має обмеження, оскільки ін’єкції дають позитивний результат лише тоді, коли риби знаходяться у стані достатньо високої стиглості. І для її досягнення необхідно створити рибі певні екологічні умови. Вважають, що два методи взаємно доповнюють один одного і сьогодні розглядаються в якості еколого-фізіологічного методу достигання статевих продуктів.
Отже, нерестовий етап розвитку має особливе значення у завершенні циклу онтогенезу і у забезпеченні єдності його різних етапів.
Зрозуміло, що теоретичні аспекти сучасного рибництва тісно пов’язані з практичною роботою відповідних напрямків рибного господарства. Виходячи з цього, доцільно визначитися і сформулювати складові, які конкретно характеризують значення теорії етапності для практики рибництва.
- Пізнання етапів розвитку риби, особливостей її біології в природних і штучних умовах дозволяє встановити потреби риби на кожному етапі розвитку і у зв’язку з цим виявити недоліки біотехніки штучного відтворення, що необхідно для раціоналізації риборозведення. Теорія етапності розвитку дозволяє біологічно обґрунтувати різні ланки біотехніки рибничих робіт і здійснити біологічний контроль за розвитком риби в природних і штучних умовах.
- Встановлення етапів розвитку в популяціях, які населяють водойму, дозволяє зрозуміти різноякісну структуру цієї популяції і різноманіття її потреб.
- Встановлення етапів розвитку промислових і малоцінних риб, які населяють одні й ті ж водойми, дозволяє вияснити, в якій степені, на яких етапах розвитку малоцінні риби становлять додаткове навантаження на кормову базу промислових риб, тобто дозволяє конкретизувати знання міжвидових відношень риб.
- Теорія етапності вимагає вивчення кормової бази з урахуванням різних споживачів на кожному етапі розвитку, тим самим конкретизує дослідження кормової бази риб. Знання особливостей перебігу кожного етапу розвитку дозволяє більш обґрунтовано прогнозувати чисельність риб в промисловому поверненні. Дефіцит певної їжі на одному з етапів розвитку може значно знижувати чисельність промислового стада.
- Зсуви термінів статевого вистигання в залежності від умов, в яких розвивається риба, може бути зареєстровано лише на основі чітких знань етапів розвитку риби.
- Знання етапності необхідно для розкриття закономірностей динаміки чисельності популяцій.
Слід зазначити, що на сьогоднішній день ще не всі питання, пов’язані з етапністю риб достатньо вивчені. Зокрема, ще не встановлені у порівняльному аспекті закономірності прояву етапності розвитку в різних екологічних групах риб. Не встановлені особливості переходів від етапу до етапу розвитку статевого вистигання з моменту викльову ембріонів із оболонки. Відсутня повна інформація щодо впливу різних умов середовища на етапи розвитку і переходу від одного етапу до другого. Важливими є встановлення еколого - морфологічних особливостей різних систем органів на різних етапах розвитку. Тобто доцільним є вивчення функцій всіх органів і їх зміни в процесі розвитку організму від зародження й до смерті. Одним з найбільш важливих питань у розвитку теорії етапності є слабо вивченою проблема переміщення з певного етапу на етап статевого вистигання риб. На особливу увагу заслуговують питання щодо впливу чинників середовища на швидкість статевого вистигання риб.
Між тим до цього часу не існує чіткої інформації щодо того, який момент в розвитку риби повинен вважатися початком періоду статевої стиглості. Окремі дослідники вважають, що такою межею слід вважати початок першого нересту, а сам період іменувати періодом розмноження. Існує також думка про те, цей період буде складатись із етапу або етапів нересту і низки інших етапів, в залежності від екологічних особливостей риби; етапу нерестової міграції, післянерестового скату, етапу виношування ембріонів, охорони нащадків тощо.
Екологічні особливості риб настільки різноманітні, що можна припустити існування у деяких з них декількох етапів розвитку в інтервалі між послідовними нерестами. У осетрових такий інтервал продовжується декілька років, протягом яких риба проходить, можливо, декілька етапів розвитку, вона суттєво змінюється, готуючись до нового розмноження. У риб з одноразовим нерестом періодом розмноження завершується існування риби. Між тим з початком останнього періоду життя зв’язані досить суттєві морфо - функціональні і екологічні зміни риби. Відомо, що початок кожного періоду розвитку співпадає з початком етапу розвитку. У тихоокеанських лососей завдяки цьому період розмноження виявляється останнім існуванням риби.
У багатьох риб протягом періоду розмноження чергуються етапи нересту і нагулу. Кожен етап нересту характеризується особливостями будови і функції риби, специфічними для екології цього етапу. Якщо нерест розтягнений в часі і в проміжках між виметуванням окремих порцій ікри біологія риби змінюється не суттєво, такий нерест об’єднується одним етапом.
Очевидно, за суворо певних умов багатократно нерестуючі риби проходять однакову кількість етапів нересту і нагулу протягом життя.
Виділення нерестового етапу у розвитку риб, вірно відображуючи біологічну своєрідність процесів розмноження, дозволяє глибше зрозуміти і деталізувати вивчення дорослих риб. Знання проходження етапів розвитку періоду і особисто періоду розмноження дозволяє поставити важливі для рибництва питання, зокрема, щодо причин затримок нересту, норм кількості нерестів у кожного виду риби за певних умов, щодо оптимальних умов, які сприяють успішному нересту, щодо прискорення цього процесу тощо.
Теорія етапності розвитку дозволяє зрозуміти пристосувальну сутність організмів в кожній момент його індивідуального життя і життя виду, виявити потреби організму на кожному етапі розвитку і тим самим дає ключ до управління цим розвитком.
В життєвому циклі риб є періоди які супроводжуються підвищеною чутливістю до зовнішніх факторів. Такі стани розглядаються в якості критичних періодів розвитку у риб, що пов’язане з дією негативних факторів впливу на певні складові життєвого циклу.
Критичними періодами розвитку є певні моменти диференцировки організму, за досягнення яких ембріони і личинки, які мають морфофізіологічні дефекти, отримані у спадку від батьків, або які виникли під впливом несприятливих умов на ембріональних і постембріональних стадіях, вже не здатні до подальшого розвитку і гинуть. Це, зазвичай, відбувається на найважливіших етапах раннього онтогенезу, особливо на початку функціонування органів, їх систем. Завдяки тому, що дефектні особини можуть мати різний характер дефектів і різну ступінь порушень органів, одні з них, з найбільш сильно вираженими дефектами, припиняють свій розвиток і гинуть раніше, що відбувається в ранні критичні періоди. Інші з меншими порушеннями, - пізніше, а треті з малими дефектами, можуть взагалі обминути критичні періоди розвитку. Але у подальшому, за несприятливих біотичних і абіотичних умов середовища, елімінація відбувається в першу чергу за рахунок цих особин. Отже, критичні періоди розвитку можна коротко назвати періодами реалізації морфо - фізіологічних дефектів, а загибель є зовнішнім проявом критичних періодів розвитку.
В критичні періоди розвитку у організмі існують свої специфічні взаємовідносини з середовищем, але вони лише до деякої міри можуть визначати ці періоди, а не в цілому.
В ембріональному і постембріональному розвиткові таких критичних періодів існує декілька. Одним із критичних періодів у багатьох видів риб є стадії кінця гаструляції і початку органогенезу, а у деяких видів - і стадія викльовування.
У деяких морських пелагофільних риб таким періодом, очевидно, можна назвати також стадії кінця дробління - бластули.
У личинковий етап розвитку виявляється також декілька критичних періодів, причому у різних груп видів риб вони припадають на різні стадії розвитку. На личинковому етапі критичні періоди не обов’язково припадають лише на кінець розсмоктування жовтка і за переходу до активного живлення.
Виходячи з проблематики її теоретичної та практичної актуальності, доцільно розглянути критичні періоди личинкового етапу окремих видів риб.
Деякі дослідники вважають, що критичним періодом у осетрових риб є перехід личинок на активне живлення. Саме в цей період реєструється масова загибель личинок за штучного відтворення представників осетрових. Це явище пов’язують з несвоєчасною годівлею і невідповідністю якості їжі потребам личинок.
Проте існує й інша точка зору, яка ґрунтується на тому, що загибель личинок досить часто спостерігається не дивлячись на сприятливі умови живлення, а крім того, вона починається ще до переходу личинок на активне живлення. Тому вважають, що загибель личинок викликана низькою якістю використаних за штучного відтворення статевих продуктів і незадовільними умовами інкубації. Підтвердженням цього висновку виявились результати спеціальних досліджень, якими встановлено, що загиблі в цей період личинки осетра характеризувались відставанням у рості за довжиною і масою, а також мали різні дефекти у будові: сутулість, вислохвостість рострума, зменшення розмірів печінки, різні порушення у будові кишківника, нерідко у поєднанні з водянкою черевної порожнини і шлунку. Все це свідчить про те, що період розвитку личинок осетрових риб перед початком і на початку активного живлення може вважатись критичним періодом розвитку. У оселедцевих критичним періодом розвитку є кінець жовткового живлення і початок активного живлення. У лососевих також чітко проявляється критичний період розвитку в момент кінцевого розсмоктування жовтка і на початку активного живлення.
Що стосується коропових риб, то у них відмічається два періоди високої смертності личинок. Перший період загибелі личинок коропа спостерігається через 5-7 днів після їх викльовування, після того, як вони повністю перейдуть на зовнішнє живлення. Загальний термін цієї загибелі - до 10-12-го дня після викльовування. Личинки, що гинули, значно відстали у рості і розвиткові від своїх ровесників, вони або зовсім не живились, або живились дуже мало. У них також відмічається відсутність складчастості слизової кишківника, або вона буває слабо розвиненою. Припускають, що в цей період відбувається реалізація дефектів у розвиткові органів травлення.
Другий період високої смертності личинок коропа, зазвичай, реєструється з 19 по 27 дні після викльовування. Ті личинки до загибелі також значно відставали у розвиткові і рості. У більшості з них не утворюється спинний плавець, а спостерігається лише скоплення на його місці мезенхімних клітин, а у деяких личинок виявляються поява лише перших променів. На відміну від них у нормальних личинок реєструють повністю сформовані плавці, за виключенням черевних. Різниця у рості інколи буває настільки високою, що крупні личинки часто заковтують дрібних або об’їдають у них хвости.
При цьому загибель личинок в період їх високої смертності не могла відбуватись із -за дефіциту відповідної їжі або від інших зовнішніх причин.
Вважають, що безпосередньою причиною загибелі личинок в період їх високої смертності є не зовнішня, а внутрішня. Цією причиною вважають низьку якість самих личинок, яка залежить у свою чергу від низької якості статевих продуктів, які продукували плідники. На виживаність личинок до деякої міри могло вплинути різке зниження або підвищення температури, що досить часто спостерігається за вирощування личинок. Отже, розглянуті два періоди високої смертності у личинок коропа можна вважати критичними періодами розвитку.
У личинок ляща також виявлено два періоди високої смертності. Перший з них припадає на 12-17-у добу, а другий - на 20-22-гу добу, які для личинок ляща є критичними.
На личинковому етапі у тарані теж знайдено, як і у ляща, 2 основних критичних періоди розвитку, з однією лише різницею, що у ляща найбільш вираженим є другий період, а у тарні - перший; крім того, у останньої тривалість періодів в середньому дещо більше.
Зовсім інші терміни періодів високої загибелі спостерігаються у личинок білого амура. Період масової загибелі личинок цього виду риб продовжується в цілому з 1-го по 6-й день після викльовування з піками, які припадають на 1-2 і дні і 4-5 дні. Експериментально встановлено, що елімінація личинок білого амура за рахунок нежиттєстійких особин відбувається в основному в період переходу на активне живлення - розсмоктування жовтка у них закінчується на 5 - 6-ий дні після викльовування.
Отже, вважають, що у личинок білого амура існує два критичних періоди розвитку. Один з них охоплює в основному перші 2 дні після вилуплення, а другий - кінець розсмоктування жовтка і початок активного живлення, тобто 4-6-й дні.
Вважають, що розвиток як ембріона в цілому так і його складових частин, маються на увазі органи і тканини, складається з деякої невеликої кількості етапів, які характеризуються певними морфо-фізіологічними ознаками. Кожні з цих етапів починаються відносно коротким критичним періодом, після якого відбуваються видимі процеси розвитку, а саме ріст і диференціація. Головною ознакою, яка характеризує критичні періоди, є висока чутливість клітин зародків до дії зовнішніх чинників, яка обумовлена значною регуляторною діяльністю в ці періоди.
Згідно сучасних уявлень періоди високої чутливості розвитку у риб виявляються у переломні моменти онтогенезу, перед найважливішими морфологічними диференцировками. Вони характеризуються високою чутливістю організму до негативного впливу зовнішніх чинників і зниженням життєстійкості організму та його загального обміну і іншими фізіологічними властивостями, які показують, що ці зміни є не лише причиною високої чутливості в дані періоди розвитку організму, але й самі є наслідком вражень клітин. Ці періоди також характеризуються посиленим диханням і зниженням темпу росту.
Різні дослідники в ембріональному розвитку риб знаходять різну кількість періодів високої чутливості - від 3 до 8, що обумовлено видоспецифічними особливостями і умовами, які формують зовнішнє середовище.
Значні морфологічні і фізіологічні дефекти, отримані личинками у спадок від батьків і в період ембріонального розвитку під впливом несприятливих умов, реалізуються, зазвичай, в критичні періоди розвитку, коли починають функціонувати певні системи органів. Критичні періоди є скоріше видовою або популяційною властивістю, а не індивідуальною, оскільки вони відносяться до популяції в цілому. В нащадках, які з’явились із ікри високої якості і за нормальних умов розвитку, критичні періоди не проявляються. Але елімінація за рахунок неякісних особин відбувається не лише в критичні періоди; за погіршення будь - яких біотичних або абіотичних умов існування у першу чергу гинуть саме вони, і така загибель, очевидно, має велике значення в динаміці виживаності поколінь.
Ступінь прояву критичних періодів залежить, з одного боку, від якісного стану організму, а з другого - від умов середовища; за несприятливих умов вони можуть проявитися більш чітко. Оскільки, як встановлено, від умов середовища залежить швидкість розвитку, то й час прояву критичних періодів до певної міри також залежить також від них. Критичні періоди в нащадках, які розвивались із статевих продуктів високої якості та за сприятливих умов середовища, не проявляються і виживаність таких нащадків достатньо висока, тому їх слід вважати популяційною властивістю, а не індивідуальною.
Критичних періодів розвитку у ранньому онтогенезі не один, а декілька, і на личинковому етапі критичні періоди не обов’язково припадають лише на кінець розсмоктування жовтка і на перехід до активного живлення.