ТЕОРЕТИЧНІ ОСНОВИ РИБНИЦТВА - І.М. Шерман - 2011
2.ТЕОРЕТИЧНІ ОСНОВИ ФОРМУВАННЯ ТА ВИКОРИСТАННЯ ПРОДУКТИВНИХ ВЛАСТИВОСТЕЙ РИБ
2.4.Еколого - фізіологічні основи природного та штучного відтворення риб
На відміну від теплокровних диких та домашніх тварин і птахів, розведення риб досить специфічне, що обумовлено їх видовою різноманітністю, яке поєднується з виключно високою значимістю абіотичних, біотичних і антропогенних факторів. При цьому перша і друга група факторів можуть впливати на риб безпосередньо, а третя група - виробничі фактори, можуть діяти як безпосередньо так і опосередковано через зміни кількісних і як наслідок, якісних характеристик умов існування, розмноження та розведення риб. Нажаль об’єктивна реальність свідчить про те, що інтенсивність впливу антропогенних факторів зберігає тенденцію підвищення, що обумовлено нарощенням господарської діяльності людини у регіональних і планетарних масштабах. Змінюється гідрологічні режими, фізико - хімічні показники континентальних і морських вод, що об'єктивно негативно впливає на видовий склад, чисельність і біомасу гідробіонтів і зокрема риб.
При цьому необхідно враховувати той факт, що критерії екологічної пластичності певного виду риб, необхідно диференціювати в зв’язку з відповідним життєвим циклами. При такому підході стає наочним, що процес розмноження риб характеризується достатньо вузькою екологічною пластичністю і є виключно консервативним, що типово для переважної більшості цінних видів риб. Розглянута біологічна особливість знаходить своє фактичне відображення в динаміці чисельності популяцій риб, коли порушується екологія розмноження, і як наслідок в окремі роки з’являються маловрожайні покоління. Так, випадіння певних видів риб із промислу відбувається протягом досить тривалого часу. Після чого їде ряд низьковрожайних поколінь молоді, причому кожне наступне покоління нижче урожайності попереднього. За умов збереження такої тенденції в часі і просторі перспектива достатньо негативна, а саме повне припинення розмноження і зникнення виду не тільки з промислу, а і складу іхтіофауни відповідної акваторії.
За таких умов рибогосподарське значення акваторії може бути збережено за умови обґрунтованих робіт з реакліматизації у поєднанні з раціональною експлуатацією водойми за принципом пасовищної аквакультури, що знайшло своє втілення в практиці сучасного світового рибництва.
Проте, згадана практика збереження рибогосподарського значення водойм, змогла стати реальністю тільки тому, що були розроблені теоретичні основи штучного риборозведення і вирощування життєстійкого рибопосадкового матеріалу який використовується в якості інтродуцентів для цілеспрямованого формування іхтіофауни природних і штучних акваторій за умови збереження туводної іхтіофауни.
Повертаючись до специфічних особливостей розмноження кожного виду доцільно розглянути цей процес в адаптаційному плані. Специфічність або видоспецифічні особливості цього процесу не що інше, як пристосування до певних умов розмноження і розвитку молоді, забезпечуючих циклічність поповнення, що необхідно для збереження виду і підтримання чисельності його популяцій в ареалі.
Чисельність поповнення, його якісні параметри залежать від якості нерестової популяції, умов ембріогенезу і раннього постембріогенезу.
Розмноження риб має свої специфічні особливості, характерні для водних тварин, і обумовлені життям у воді, що необхідно враховувати при розведенні. Одночасно з цим, на відміну від теплокровних тварин, що мешкають у воді, у абсолютної більшості риб запліднення яйцеклітин проходить поза межами материнського організму, а саме в зовнішньому середовищі. Яйцеклітини і сперматозоїди до настання запліднення певний час знаходяться у воді, поза особинами - плідниками, де і відбувається проникнення сперматозоїдів в яйцеклітину і формування зиготи, що і свідчить про факт запліднення.
Виходячи з викладеного, зрозуміло, що фактори середовища безпосередньо впливають на процес розмноження в природних умовах, маючи провідне значення в процесі розведення риб. При цьому доцільно пам’ятати, що нерест є виключно ваговою складовою відтворення, але це своєрідний фінал, якому передує процес формування відтворної системи. Тривалість цього процесу і якість статевих продуктів залежить не тільки від видоспецифічних особливостей, тут вагоме значення набувають абіотичні і біотичні фактори середовища, рівень їх залежності від антропогенних чинників.
Для одержання повноцінних статевих продуктів, які формуються у відповідних статевих залозах, необхідно володіти інформацією відносно процесу, перебіг якого відбувається в статевих залозах до досягнення статевої стиглості.
І стадія - статевозрілі (ювенальні) молоді особини. Статеві залози мають вигляд тонких прозорих тяжів, які прилягають до стінок порожнини тіла. Статеві клітини самиць можуть бути представлені оогоніями, або молодими овоцитами періоду протоплазматичного росту, статеві клітини самців представлені сперматогоніями.
Овогонії, або первинні статеві клітини самиць утворюються із зачаткового епітелію протягом всього життя. Ці клітини округлої форми і невеликих розмірів, що не дає можливості виявити їх візуально. Для цього виготовляють гістологічні препарати і за великого збільшення розглядають під мікроскопом. Вони прозорі, мають відносно крупне ядро, оточене тонким шаром протоплазми (рис. 2.1., 2.2).

а б
Рис. 2.1. - Ділянки яєчників 1 (а) і II (б) стадії зрілості:
1 - овогонії; 2 - ооцити початку протоплазматичного росту; 3 - ооцити кінця протоплазматичного росту

Рис. 2.2. - Ооцити з яєчииків III (а) і IV (б) стадії зрілості: 1 - ядро; 2 - каплі жиру 3 - жовток, 4 вакуолі; 5 - оболонка ооцита.
Овогонії розвиваються і діляться, що веде до збільшення їх кількості. Цей період в розвитку статевих клітин отримав назву періоду поділу овогоній. Після цього частина овогоній припиняє процес поділу, проходить період перетворень в ядрі (так званого сенаптенного шляху) і починають збільшуватись в розмірах (ріст). Статеві клітини періоду росту мають назву овоцитів. Ріст молодих овоцитів відбувається за рахунок збільшення кількості протоплазми — цей період їх розвитку називають періодом протоплазматичного, або малого росту (2.3).
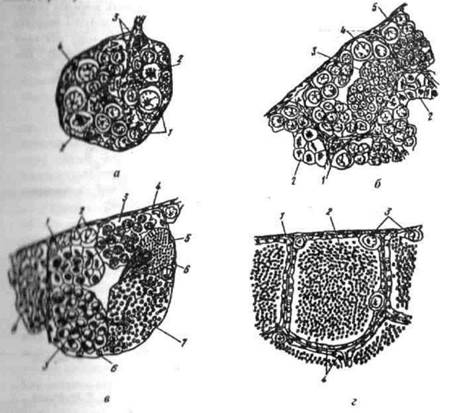
Рис. 2.3. - Поперечний розріз судин сім'яників на різних стадіях зрілості ( а -1; б - II; в - III; г - IV стадії ): 1 - Кровеносна судина з еритроцитами; 2 - сперматогонія; 3 - поділ сперматогонії; 4 - оболонка сім’яника; 5 - циста з сперматоцитами І порядку; б - циста із зрілими сперматоцитами; 7 - сперматозоїди; 8 - циста із сперматидами; 9 - циста з поділеними сперматоцитами II порядку; 10 - фолікулярний епітелій
Сперматогонії — первинні статеві клітини самців, які утворюються із перитонеального епітелію. Вони присутні у самців в будь-який період року і на будь-якій стадії статевого циклу. Це найбільші клітини в сім'яниках, для них характерна округла форма і відносно крупне ядро, оточене товстим шаром цитоплазми.
ІІ стадія — дозріваючі особини або особини, в яких проходить розвиток статевих продуктів після нересту. Яєчники прозорі, практично безбарвні. Вздовж проходить велика кровоносна судина, що має відгалуження. В яєчниках деяких риб, зокрема в осетрових, наявна значна кількість жирової тканини. Під час розгляду яєчників при збільшенні добре розрізняються овоцити періоду протоплазматичного росту, які складають основну масу статевих клітин. Значна кількість овоцитів проходить кінцеві фази цього періоду. Вони мають відносно великі розміри за рахунок збільшення ядра і об'єму цитоплазми (2.4).
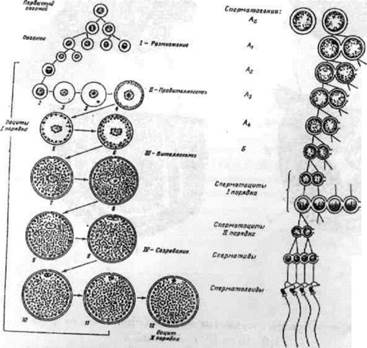
Рис. - 2.4. - Схема овогенезу і сперматогенезу
Окремі овоцити закінчили протоплазматичний ріст і їх уже можна розрізнити неозброєним оком. Навколо овоцитів проходить закладка шару фолікулярних клітин, які утворюються із зародкового епітелія яєчників. Це початок формування фолікулярної оболонки. Паралельно з овоцитами, що пройшли період протоплазматичного росту, (в яєчниках є також овогонії і овоцити початкових фаз періоду малого росту. Ці статеві клітини виступають як резервний фонд, який може бути використаний організмом самок поліциклічних риб (багаторазово нерестуючі протягом життя) після майбутнього нересту в черговому циклі дозрівання статевих продуктів.
Сім'яники представлені округлими тяжами або тонкими нитками сіруватого або біло-рожевого кольору. У прохідних оселедців, лососів та у деяких інших видів риб через сильний розвиток кровоносних судин сім'яники можуть набувати різних відтінків від рожевого до багряно- червоного. Статеві клітини самців представлені сперматогоніями в стані розмноження.
Сперматогонії декілька разів поділяються, при цьому збільшується їх кількість і зменшуються розміри. Таким чином, з кожної початкової крупної сперматогонії формується група більш дрібних сперматогоній, що мають спільну оболонку. Ці групи статевих клітин мають назву цист. Розмноження сперматогоній приводить до збільшення розмірів сім'яника, що супроводжується його помутнінням і втратою прозорості.
ІІІ стадія — статеві залози добре розвинені. Яєчники заповнюють половину об'єму черевної порожнини і містять овоцити, видимі неозброєним оком. Овоцити збільшують свої розміри за рахунок зростання об'єму протоплазми і в результаті накопичення трофічних речовин, представлених зернами (гранулами) жовтка і краплинами жиру. Цей період розвитку овоцитів має назву періоду трофоплазматичного, або великого росту. Завершення періоду росту характеризується тим, що овоцити досягають розмірів, які в багато десятків разів перевищують початкові розміри овогоній. Овоцити при цьому втрачають прозорість, мутніють і набувають за рахунок жирових капель і гранул жовтка забарвлення від світло-жовтого з різними відтінками до яскраво-жовтогарячого, що приводить до загальної зміни кольору яєчників. У осетрових завдяки акумуляції у верхньому шарі цитоплазми дрібних гранул бурувато-чорного пігменту овоцити набувають характерного для них темного забарвлення. Крім крапель жиру і гранул жовтка, в овоцитах костистих риб з'являються вакуолі, що містять речовини вуглеводної природи.
На цій стадії проходить формування оболонки овоциту. Спочатку утворюються мікролійки на поверхні овоциту, біля основи яких формується тонкий шар гомогенного, не маючого структури матеріалу, безпосередньо контактуючого з поверхнею овоциту і на деякій відстані від фолікулярних клітин. При збільшенні накопичення жовткових включень в овоциті формується додатковий шар, що складається із пучків трубчастих структурних елементів. Він розмішується безпосередньо між тонким гомогенним шаром і поверхнею цитоплазми. Після цього внутрішній структурний шар переходить в гомогенний зовнішній і обидва шари утворюють єдину оболонку. У осетрових, кети, салаки, окуня, атлантичних і тихоокеанських оселедців, деяких інших видів риб ця оболонка складається з двох шарів, зовнішнього і внутрішнього. У багатьох інших видів вона, як правило, має тільки один шар. Оболонка пронизана канальцями, в яких лежать ворсинки овоциту. Під мікроскопом на оболонці овоциту видно характерну радіальну почерканість, що утворена найтоншими порами канальців, що і лежить в основі назви — радіально-почеркана оболонка (zona radiata). Сама оболонка представлена міжклітинною речовиною, яка пронизана радіальними канальцями, в яких проходять мікроворсинки овоциту. Таким чином овоцит зі сформованою zona radiata оточений фолікулярними клітинами, які утворюють фолікулярну оболонку, або фолікулу. Зовнішня поверхня фолікули вкрита сполучною оболонкою. Над zona radiata в овоциті ряду видів риб формується іще зовнішня оболонка, яка буває представлена одним шаром гомогенної або ворсинчастої будови. При цьому у різних видів риб відстежуються специфічні особливості. Плітка має ворсисту зовнішню оболонку, у осетрових на поверхні zona radiata утворюється друга оболонка, яка складається із радіально розташованих стовпчиків (щільниковий шар). Таку оболонку називають ніздрюватою, драглистою або щільниковою. Для ряду риб характерне утворення наростів на zona radiata у вигляді гребенів і ниток. У одних видів риб ці нарости розташовані рівномірно по всій zona radiata, а в інших вони розміщуються на локальних ділянках. Особливості будови оболонок овоцитів різних видів риб обумовлені адаптацією ікринки до умов ембріогенезу, які виробились в процесі філогенезу. Так у риб, ікра яких розвивається в товщі води, можливість механічного травмування мінімальна і потрібне максимальне
полегшення ікринок для гарної їх плавучості, овоцити мають дуже просту будову оболонок, представлених лише zona radiata, або додаткове покриття зверху дуже тонким гомогенним шаром. У риб, які відкладають ікру на грунт і занурені рослини, де вірогідність механічного пошкодження досить висока, овоцити мають zona radiata і зовнішню оболонку, якщо ж її немає, то на zona radiata існують нарости. Коли яйцеклітина попадає у воду, зовнішня оболонка або нарости набрякають, стають клейкими і виступають в якості специфічного органу, який забезпечує фіксацію ікри на субстраті.
Очевидно, що для третьої стадії стиглості яєчників характерна наявність овоцитів періоду трофоплазматичного росту. Поряд з цим в яєчниках поліциклічних риб присутні статеві клітини резервного фонду, представлені овоцитами і овоцитами періоду протоплазматичного росту.
Сім'яники третьої стадії стиглості значно збільшуються в об'ємі, стають щільними і тугими. На початку стадії їх колір рожево-сірий, а в кінці жовтувато-білий. Сім'яні канальці заповнені цистами і статевими клітинами. Сперматогонії, пройшовши період розмноження на другій стадії стиглості сім'яників, вступають в період росту і трансформуються в сперматоцити першого порядку. Спостерігається деяке збільшення розмірів цих клітин, що супроводжується складними ядерними перетвореннями, які відіграють досить значну роль в спадковості. Після цього чоловічі статеві клітини вступають в період достигання і двічі послідовно діляться. Наслідком поділу є утворення із кожного сперматозоїда першого порядку двох сперматозоїдів другого порядку, а потім чотирьох сперматозоїдів, які відрізняються від початкових меншими розмірами, відносно великим ядром, яке оточене найтоншим шаром цитоплазми. Внаслідок двократного поділу кількість хромосом в сперматидах зменшується в два рази, що тісно пов'язане зі спадковістю. Сперматиди, що утворились, вступають в період формування і поступово перетворюються в зрілі сперматозоїди. Форма головки сперматозоїдів у різних видів риб не однакова. У більшості костистих риб форма головки сперматозоїда кульовидна, яйцевидна, жолудеподібна, у осетрових і у деяких інших видів форма головки паличковидна (рис.2.5).
Основна маса головки сперматозоїда представлена ядром, у осетрових в передній частині головки знаходиться невелике утворення — акросома, формою нагадуючи шапочку, у інших видів аналогічне утворення відсутнє. До задньої частини голівки прилягає середня частина сперматозоїда, в утворенні якої беруть участь клітинний центр, мітохондрії і сперматиди. Середня частина, в свою чергу, переходить в хвіст, утворений основною ниткою, яка оточена шаром цитоплазми, і вкритий на поверхні плазматичною мембраною. Хвіст забезпечує рухові функції сперматозоїда. Після завершення процесу утворення в цистах сперматозоїдів оболонка цист руйнується, і вони вільно лежать в сім'яному канальні. Сперматозоїди нерухомі в сперміальній рідині і набувають здатності руху тільки за контакту з водою.
Це найменші клітини із усіх вихідних статевих клітин, що продукуються сім'яниками.
Підсумовуючи викладене, необхідно акцентувати увагу на тому, що в сім'яниках третьої стадії стиглості спочатку є сперматогонії, сперматоцити першого і другого порядків та сперматиди, а при завершенні стадії з'являються окремі групи зрілих сперматозоїдів.
IV стадія — ікринки великі, практично повністю досягли відповідних розмірів і маси і займають більшу частину порожнини тіла. Колір яєчників у різних видів варіює в широких межах. Зазвичай він жовто-жовтогарячий, а в осетрових риб сірий, або практично чорний, що пов'язано зі ступенем пігментації овоцитів. Навіть у самок одного виду риб колір яєчників може бути різним. В яєчниках є овоцити, які закінчили трофоплазматичний ріст і готові до овуляції при ікрометанні в період нересту. В яєчниках розглядаємої стадії деяких видів риб поряд з цими статевими клітинами є також овогонії і овоцити протоплазматичного росту, що складають своєрідний резервний фонд. Овоцити, які закінчили трофоплазматичний ріст і досягли характерних розмірів для ікринок даного виду риб, вступають в період достигання, а саме в період, що завершує розвиток статевих клітин. Характерною діагностичною ознакою цього явища є зміщення ядра овоцита до мікропіле. Костисті риби мають одне мікропіле, в осетрових мікропіле декілька. Мікропіле являє собою канал, який пронизує zona radiata, а також драглисту оболонку, якщо вона є в овоцита. Цей канал має форму лійкоподібного заглиблення на поверхні оболонки (оболонок), яке закінчується коротким кінцевим канальцем, що відкривається в цитоплазму в середині zona radiata. Зміщення ядра в овоциті приводить до полярності розміщення ядра і жовтка, на одному полюсі — анімальному — ядро і основна маса цитоплазми, на другому — вегетативному — жовток. В подальшому повне або часткове злиття жовтка з жиром надає прозорості овоцитам (рис. 2.6.).
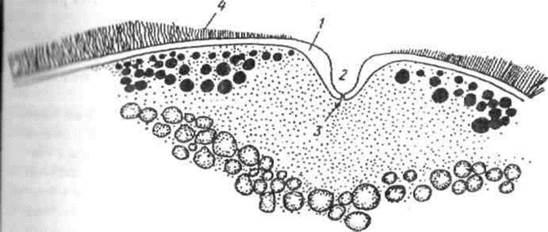
Рис. 2.6. - Мікропіле зрілого ооциту великоротого буфало:
1 - воронка ооциту; 2 - воронка мікропіле; 3 - каналець мікропіле; 4 - хоріон.
Сім'яники на четвертій стадії зрілості досягають найбільшої величини, набувають молочно-білого кольору, і в них завершується спарматогенез В сім'яних канальцях статевих залоз містяться зрілі сперматозоїди, що вийшли з цист. При цьому в сім'яниках зберігається запасний фонд статевих клітин, представлених сперматогоніями. При пальпуванні черевця риби із генітального отвору з'являються краплини сперми, що мають консистенцію густої сметани.
V стадія — статеві продукти текучі і йде нерест риб. При обережному пальпуванні черевця риби ікра і сперма вільно витікають із генітального отвору. Цією стадією завершується процес підготовки овоцитів до запліднення. В овоцитів осетрових риб ядерця ядра розчиняються, а його оболонка набуває складчастості на вегетативній стороні, через яку каріоплазма частково виходить в цитоплазму. В цій частині овоциту утворюється велика лакуна, яка заповнена матеріалом, що не містить жовткових гранул. Ядро стрімко скорочується в розмірі, плазма в значній мірі змішується з цитоплазмою, а незначна її частина представлена острівцями, що збереглися, і які створюють своєрідну розлогу сітку. Після завершення цього процесу — оболонка ядра розчиняється і розпочинається мейотичний поділ. Процес завершується звільненням овоцитів від фолікулярної і сполучнотканинної оболонок. Фолікули розриваються, і овоцити у більшості видів риб випадають в порожнину яєчників, а в лососевих і осетрових, котрі мають незамкнені яєчники — в порожнину тіла.
Розрив фолікул і вихід в порожнину яєчників або тіла (овуляція) у одних видів риб протікає синхронно, а в інших цей процес розтягнутий в часі. Фолікули, що лопнули, залишаються в яєчниках, де з часом вони резорбуються. В яєчниках поліциклічних риб окрім тріснувших фолікул, містяться статеві клітини резервного фонду.
На п'ятій стадії стиглості в сім'яниках утворюється сім'яна рідина, яка розбавляє концентрацію сперматозоїдів і викликає їх вільне витікання. Сперма має консистенцію молока або рідкої сметани, сім'яники м'які, і по мірі витікання сперми їх розмір зменшується.
Стадія VI - II — відображує післянерестовий стан статевих залоз, яйцеклітини і сперматозоїди, виведені в процесі нересту. В цей період яєчники мають невеликі розміри, характеризуються дряблістю і запаленнями. Типовим є багряно-червоний колір, викликаний крововиливами, пов'язаними з розривами фолікул. Спорожнілі фолікули і окремі зрілі ікринки, що залишились в яєчниках не виведеними, в процесі нересту резорбуються. Для поліциклічних риб характерна наявність в яєчниках резерву, представленого недозрілими статевими клітинами. Таке явище не спостерігається у моноциклічних риб, нерестуючих тільки раз в житті. Склад статевих клітин, що утворюють резервний фонд поліциклічних видів риб, поданий структурами, які відповідають другій стадії стиглості яєчників. Таким чином, черговий цикл в цьому випадку починається з другої стадії стиглості яєчників (рис. 2.7.).
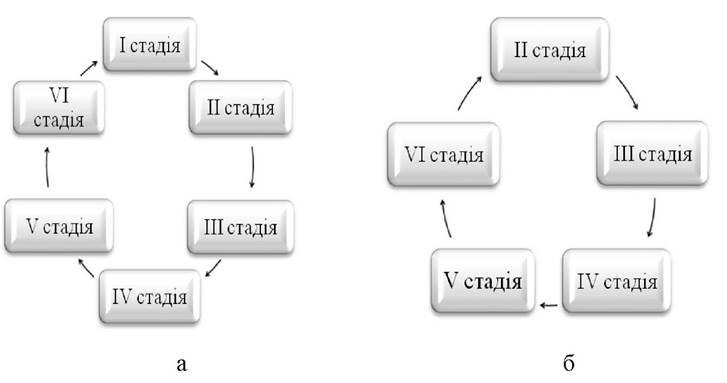
Рис. 2.7. - Стадії розвитку статевих залоз риб а - моноциклічних та першонерестуючих; б - поліциклічних та багатократнонерестуючих риб.
Зрозуміло, що за умов нормального проходження відповідних стадій статевої стиглості залоз будуть отримані високоякісні статеві продукти.
При цьому виключне значення набувають абіотичні фактори середовища, особливо його окремі параметри, які забезпечують в природних умовах нормальне живлення, а в штучних умовах цей процес може корегуватися кормами.
Розглядаючи еколого - фізіологічні основи природного та штучного відтворення риб, необхідно наголосити на тому, що в основі лежить біологія розмноження, яка дуже складна, видоспецифічна регуляція якої, має нейрогуморальний характер. При цьому існують дві функціональні складові - наявність статевостиглих самиць і самців з одного боку і відповідність умов для нересту, які передбачають суму факторів абіотичного і біотичного походження.
В складі іхтіофауни є види, які віднесені до еврибіонтів, саме вони переважно легко адаптуються до різних умов відтворення, але переважна більшість цінних об’єктів з господарських позицій є стенобіонтами, вони легко втрачають здатність до відтворення на тлі зміни абіотичних і
біотичних факторів середовища.
В цьому зв’язку історія питання полягає в тому, що фахівці з давніх- давен, в процесі доместикації різних видів риб зіткнулися з ситуацією різного рівня здатності до відтворення в штучних умовах, що було важливим при виборі видів, які доцільно і можливо культивувати.
Протягом тривалого часу фахівці спрямовували свої зусилля на створення умов максимально наближених до природних з метою отримання ефекту відтворення в штучних умовах. На цьому шляху були досягненні певні успіхи, але для багатьох бажаних видів риб проблема залишилася далекою від вирішення, що знайшло своє відображення в концепції екологічних методів витримування плідників для забезпечення статевої стиглості. Тривалі дослідження ефективності екологічних методів стимуляції і достигання статевих залоз показало обмеженість цього напрямку, а у роботі з певними видами риб повну непридатність в умовах рибничих господарств.
В 30 - 40 роках ХХ століття сумісні зусилля фахівців в галузі фізіології, іхтіології, рибництва дозволили суттєво розширити уявлення відносно механізму нересту риб в природних умовах, що відкрило нові можливості для культивування риб в режимі аквакультури.
Сутність цього механізму полягає в тому, що плідники певних видів риб, які мають статеві залози у завершеній IV стадії стиглості при настанні нерестових температур в природних умовах починають рух у напряму нерестовищ. На цьому шляху поступово відбувається перехід з IV стадії в V стадію стиглості. Але завершеність цієї стадії відбувається безумовно тільки на нерестовищах за умови наявності всіх відповідних факторів абіотичного і біотичного походження.
Сума відповідних факторів за якісними і кількісними характеристиками сприймається органами чуття риб, які передають відповідні сигнали у головний мозок, де вони досягають критичної маси і примушують головний мозок впливати на відповідну залозу внутрішньої секреції. Під дією відповідного сигналу головного мозку гіпофіз, який і є цією залозою, виділяє або викидає у кров попередньо напрацьований гонадотропний гормон. Гонадотропний гормон, рухаючись у складі крові досягає статевих залоз статевостиглих особин, що забезпечує перехід від завершеної IV в V стадію стиглості. Наочним критерієм цього явища є вільне витікання відповідно ікри і молок, що при їх безпосередньому контакті забезпечує процес осімінення.
Безпосередньо після осімінення сперматозоїди активовані водою проникають в яйцеклітину, утворюється зигота - початок ембріогенезу.
В штучних умовах утримання ряд видів - стенобіонтів можуть не досягати завершеної IV стадії стиглості, а ряд видів досягає цієї стадії, що залежить від умов утримання. Поряд з цим перехід у V стадію достатньо проблематичний і не кожний вид демонструє таку здатність. Використання максимальної можливості екологізації умов утримання в режимі аквакультури не дає відповідних гарантій для ряду видів риб, а певні види практично не реагують на такі заходи, перехід з завершеної IV стадії стиглості в V стадію не відбувається, що виключає можливість отримання повноцінних статевих продуктів.
Відсутність можливості нересту в природних умовах відповідних видів риб полягає в тому, що органи чуття не мають можливості отримати відповідне подразнення, в головний мозок не надходять відповідні сигнали і він не віддає своєрідну команду гіпофізу. У свою чергу гіпофіз, як залоза внутрішньої секреції, залишається пасивним, не викидає в кров гонадотропний гормон, який, насичуючи кров, впливає на перехід статевих залоз з завершеної IV в V стадію стиглості. Ікра і молоки не виштовхуються в зовнішнє середовище, що виключає можливість відтворення в штучних умовах.
Засвоєння попередньо викладеної інформації дозволяє констатувати - без відповідної концентрації гонадотропного гормону гіпофізу у крові риб перехід статевих продуктів із завершеної IV стадії стиглості в V стадію стиглості, коли статеві продукти вільно витікають у зовнішнє середовище, принципово не є можливим.
Концептуальний розгляд об'єктивно існуючого механізму нересту приводить до висновку - для забезпечення ефекту відтворення необхідно досягти оптимальної концентрації гонадотропного гормону гіпофізу у складі крові статевозрілих особин, що дозволяє перевести завершену IV стадію стиглості статевих залоз в V стадію стиглості, яка забезпечує вільне витікання у зовнішнє середовище статевих продуктів. У свою чергу їхній контакт між собою розглядається як осімінення, за умови якого виникає вірогідність потенційного запліднення, що у свою чергу вимагає відповідних умов і демонструє різну ефективність.
Керуючись викладеним, були запропоновані, так звані гіпофізарні ін’єкції препаратами природного походження, отриманих від статевозрілих особин певного виду, вилученого з природних умов. Рухаючись по цьому шляху, вчені і практики прийшли до можливості використання препарату від інших видів риб. Як в першому, так і в другому варіантах, гормон гіпофізу можна отримувати виключно у статевозрілих особин і бажано у преднерестовий період.
Розглядаючи можливість отримання ефекту нересту в штучних умовах, і спираючись на тенденцію екологізації умов отримання плідників у преднерестовий період в режимі культивування, цей метод отримав назву екологічного. Об’єктивна оцінка цього методу свідчить про його суттєву обмеженість, що є перепоною на шляху до промислового використання.
Розглядаючи можливість отримання ефекту нересту в штучних умовах за рахунок штучного введення препарату гіпофізу і надходження його в кров статевостиглих особин, далеко не у всіх випадках, не у всіх видів риб спостерігався позитивний результат.
Для риб, як для пойкілотермних тварин, спусковим механізмом для початку перед нерестової міграції, яка переходить у нерестову, є температура води. Для теплолюбивих видів риб, з весняно - літнім нерестом, це підвищення температури води і наближення її до нерестових температур. Для холодолюбивих видів риб, з осінньо - зимовим нерестом, це зниження температури води і наближення до нерестових температур.
При відповідності термічного режиму вимогам виду, за наявністю ряду інших абіотичних факторів, в межах норм для виду, дія штучно введеного гонадотропного гормону гіпофізу адекватна і забезпечує перехід статевих залоз в V стадію стиглості.
Таким чином стало зрозуміло, що ефект дії гонадотропного гормону гіпофізу за його штучного введення забезпечує позитивний ефект виключно за наявності певних (часткових, але необхідних) абіотичних параметрів середовища, серед яких домінує температура води.
Виходячи з запропонованої інформації, стало зрозуміло, що ефективність фізіологічної складової може дати позитивний ефект виключно за умов відповідності певних абіотичних параметрів середовища.
Саме тому вважаємо, що існуючий термін комбінований метод доцільно замінити на еколого - фізіологічний метод стимуляції переходу статевих залоз риб з IV в V з метою отримання статевих продуктів високої якості в штучних умовах.
Сьогодні поряд з гонадотропним гормоном природного походження широко розповсюджені синтезовані препарати аналогічної дії. Таке розповсюдження стимуляторів дозволило використовувати оптимальні умови для відповідних видів риб, а сам метод отримав широке розповсюдження у практиці сучасного рибництва.
В практичній роботі з розведення риб важливими складовими є знання теорії запліднення у риб, вибірковості запліднення, впливу якості сперміїв на формування потомства, особливостей підбору плідників для забезпечення одержання якісного потомства, вплив віку плідників на якісні показники розведення.
Розглядаючи теорію запліднення риб, доцільно запропонувати коротку історичну інформацію, що проливає світло на еволюцію поглядів, які пов’язані з розумінням самої суті запліднення.
Тут необхідно відзначити, що у вісімнадцятому столітті вчені не мали чіткої уяви про процеси розмноження тварин, що населяють водойми. Навіть найвидатніші натуралісти того часу помилялись відносно ролі сперматозоїдів в заплідненні яйцеклітини, вважали їх сторонньою, несуттєво важливою частиною статевих клітин самців.
В своїх працях С.Л. Якобі вирішив встановити, чи не відкладають риби ікру, яка вже була раніше запліднена в результаті парування, аналогічно з птахами, які відкладають запліднені яйця. Для відповіді на це запитання він, працюючи з фореллю, утримував самок і самців окремо, що дозволило зібрати ікру, відкладену самками за відсутності самців. Цю ікру переносили у воду, де були створені всі необхідні умови для нормального розвитку заплідненої ікри. Проте неодноразове повторення цього експерименту протягом декількох років не дозволило отримати нащадків із ікри, яку брали від самок у відсутності самців. Це дозволило досліднику переконатися в тому, що запліднення у риб відбувається поза материнським організмом. Це на сьогодні абсолютно очевидне явище в свій час було вирішальним у визначенні шляхів підходу до вирішення проблеми штучного риборозведення.
Отримавши якісно нову інформацію, значення якої важко переоцінити, С.Л. Якобі продовжив дослідження, спрямувавши свої зусилля на встановлення місць і особливостей запліднення у риб в природних умовах.
Багато років спостерігаючи за природним відтворенням форелі, дослідник установив, що в листопаді-лютому самці і самки збираються в зграї і локалізуються на мілководних ділянках струмків. Для місць концентрації форелі характерні швидка течія і вистелене гравієм дно струмка. При цьому особини двох статей, штовхаючи черевцями гравій, роблять в ньому заглиблення, куди самки відкладають ікру, а самці виділяють молочко, «так як в кожній краплі сім'я міститься багато сот «анімалькулей» (сперматозоїдів), то вода такого струмка в тих місцях і в час, коли нерестують форелі, наповнені ними. Звідси не дивно, що в кожне яйце проникає анімалькуль і запліднює його». Виклавши свої думки таким чином, С.Л. Якобі не залишив нащадкам сумніву відносно того, що він об'єктивно висловив і чітко уявив суть процесу запліднення ікри риб в природних умовах. Більш того, досягнуті результати привели його до оригінальної думки відтворити процес запліднення риб в посудині з водою, де в результаті змішування ікри і молочка відбувалось запліднення.
Отримані результати фактично стали одним із способів штучного осіменіння ікри, який забезпечує за інших рівних умов високий відсоток заплідненості і успішно використовується до цього часу.
Окрилений результатами досліджень, С.Л. Якобі розширив масштаби робіт. Працюючи з молочком і ікрою лососів, інших риб, він супроводжував експерименти багаторазовими повторами, проте незмінно здобувались аналогічні результати, що виключало сумніви відносно їх достовірності. Таким чином, в штучних умовах був відтворений процес, що відбувається в природних умовах, чим була доведена принципова можливість штучного осіменіння ікри риб, яке в подальшому забезпечує запліднення.
Теоретичні дослідження видатних натуралістів дев'ятнадцятого століття мали виключне значення в галузі пізнань процесу запліднення, заклавши основи сучасної ембріології. В цьому зв'язку, узагальнення провідних вчених - «всіляке дозріле яйце, яке прийшло в контакт з сім'яною рідиною тварини того ж виду, запліднюється, і в результаті відбувається розвиток зародка, якщо яйце знаходиться в сприятливих умовах» є проникливим. Розвиваючи і узагальнюючи раніше сформульовану концепцію, К.М.Бер пише: «В усякому випадку ясно, що всі яйця, які призначені природою для запліднення і розвитку поза материнським тілом, дуже легко можуть бути запліднені штучним шляхом». Очевидну зацікавленість і зауваження має К.М.Бер про те, що термін «штучне запліднення» застосовується вченими в значенні штучного нанесення рідини чоловічих статевих залоз на яйце, обминаючи статевий акт. Таким чином очевидно, що існує принципова різниця між осіменінням і заплідненням, незалежно від того, чи явище проходить в природних чи штучних умовах.
В усякому випадку осіменіння може бути як природним, так і штучним, а запліднення — тільки природним.
В зв'язку з заплідненням ікри риб в штучних умовах велику зацікавленість представляють спостереження відомих фахівців в галузі штучного розведення риб, які в процесі роботи із штучного розведення миньків емпіричним шляхом встановили, що незапліднена ікра відрізняється від заплідненої рихлістю, помутнінням і втратою клейкості, але це не позначається на її розмірах. Це спостереження підтвердило інформацію і надало практичному риборозведенню критерії якості ікри.
Видатний рибовод В.П. Врасський, починаючи з 1854 року, присвятивши своє життя актуальним проблемам риборозведення, поряд з виключно значними фундаментальними дослідженнями, також багато зробив в галузі штучного розведення риб, об'єктивно оцінивши його практичне значення, поставив собі завдання «щоб там не сталося розкрити таємниці і удосконалити прийоми штучного запліднення». Не менш цікаві його практичні висновки, що не втратили актуальності і на сьогодні, відносно здатності ікринок бути заплідненими. «Якщо при витискуванні ікри не виходить разом з нею кров і вона виходить легко, то можна бути впевненими, що виходить цілком зріла, нормально відокремлена від яєчника ікра, і якщо ще не всі ікринки дозріли, то незрілі не вийдуть». В.П. Врасський рахував, «що необхідно приписувати невдачу в заплідненні якій-небудь іншій причині», а не тому, що самки віддають ікру, яка не повністю дозріла.
Підсумовуючи свої дослідження в існуючій на той час спеціальній літературі матеріали, В.П. Врасський пише, що ікра лежить в порожнині черева в нейтральному (не активному) стані і що нейтральний стан ікри зараз же по виході її звідти у воду припиняється; тут з нею відбуваються самі очевидні зміни: вона починає всмоктувати в себе воду, через що набрякає її зовнішня оболонка і стає значно товщою і твердішою... Коли закінчилося це всмоктування, живчики не можуть проникнути в яйце...». В зв'язку з можливістю запліднення дослідник пише, що сперматозоїди можуть знаходитися в сім'яниках, за певних умов, протягом досить тривалого часу, «але... будучи вийняті із риби і змішані з водою, в перші миттєвості після змішування, хоча вони і чинять під мікроскопом самий бадьорий рух, але він скоро до того ослаблюється, що вже по закінченні 1-2 хвилин тільки в деяких із них помітні судорожні, так би мовити, передсмертні рухи, які ще досить довго продовжуються».
Дійшовши висновку про те, що ефективний процес запліднення можливий тільки за умови мінімального проміжку в часі між народженням у воді яйцеклітин, сперматозоїдів і їх контакту між собою, В.П. Врасський пише «...я пересвідчився, що чим більшим був проміжок часу між випусканням ікри і молочка у воду і змішування їх між собою, тим менше ікринок запліднювалось».
Дослідження В.П. Врасського мали величезне теоретичне значення і зробили великий внесок не тільки в пізнання принципів запліднення риб, але і заклали основи сучасного риборозведення. Значна частина встановлених ним принципів покладена в основу сучасних методів штучного розведення багатьох видів риб.
У зв'язку з загальною проблемою розведення риб необхідно зауважити, що в сучасній рибогосподарській літературі спостерігаються випадки подвійного уявлення і тлумачення суті деяких термінів, а в окремих публікаціях ми стикаємось навіть з помилковою трактовкою спеціальних термінів. В цьому плані це, в першу чергу, стосується таких понять, як осіменіння і запліднення, що зумовлює необхідність їх розтлумачення, інакше різко погіршується наступне сприйняття матеріалів даного розділу.
Осіменіння — це стикання, контакт сперми з ікрою (сперматозоїда з яйцеклітиною). Суть цього процесу залишається незмінною, вона не залежить від того, чи сперму виділяє самець на ікру, яка викинута самкою в умовах природної водойми, чи рибовод в умовах штучного риборозведення забезпечує контакт яйцеклітини і сперматозоїдів. Таким чином осіменіння може бути натуральним і штучним.
Запліднення — це злиття жіночих і чоловічих статевих клітин, що дає початок новій якості — зиготі, яка в свою чергу дає початок новому життю.
Цей процес не може бути штучним. Це природний процес, суть якого зводиться до наступного: сперматозоїд через мікропіле проникає всередину яйцеклітини, де відбувається злиття чоловічих і жіночих статевих клітин. В результаті цього утворюється зигота, яка несе в собі чоловічий і жіночий початок. Вона шляхом складних поділів в процесі ембріогенезу перетворюється в багатоклітинний зародок, або ембріон.
Потрапивши у воду, ікринки досить швидко втрачають здатність до запліднення. Для більшості видів риб цей період не перевищує 2-3 хвилин, а іноді і того менше. Для сперміїв риб характерна відсутність виявленого таксису, або іншими словами вони не здатні до активного пошуку яйцеклітин. Ефективність стикання сперміїв з яйцеклітинами обумовлена не їх індивідуальною активністю, а чисельністю і рухливістю. При контакті з водою рухова активність сперміїв стрімко зростає, максимум спостерігається через 50-150 секунд, а потім поступово знижується. В.А.Амінєва і А.А.Яржомбек пропонують фізіологічне бачення процесу запліднення. Такий підхід, на нашу думку, дозволяє значно краще сприймати дане явище і буде сприяти розширенню загальноприйнятого уявлення дещо нетрадиційно.
За свій короткий період життя сперматозоїд може подолати відстань, що не перевищує 20-100 його власних довжин, тобто не більше 1 сантиметру. Як відомо, процес проникнення сперматозоїда в яйцеклітину здійснюється через мікропіле - характерний отвір в зовнішній оболонці, що має лійковидну форму.
Поряд з мікропіле для ікри костистих риб типова наявність великої кількості дрібних rop, пронизуючих її наскрізь. На розрізі ці пори сприймаються як покреслєння, тому оболонка ікринки має і іншу назву — промениста оболонка. Пори променистої оболонки легко проникні для молекул води, але виключають проникнення макромолекул. Перивітеліновий простір в ікрі утворюється незалежно від запліднення. Вода проникає під оболонку ікри і викликає набрякання кортикальних (коркових) альвеол, особливих пухирців, заповнених колоїдом. Альвеоли розриваються, колоїд набрякає і збільшується в об'ємі, розтягуючи еластичну в цей час променисту оболонку. Під нею утворюється перивітеліновий простір, який функціонально захищає зародок від механічних пошкоджень. Утворившись, перивітеліновий простір дає можливість проникнення сперматозоїда, а звідси і запліднення.
Ікра, яка овулювала, може деякий час зберігати здатність до запліднення, якщо її тримати прямо в тілі виловленої риби або в оваріальній рідині. Ікра форелей і лососів може зберегти здатність до запліднення декілька діб за нульової температури, ікра сигових і коропових — декілька годин.
З цього можна зробити висновок, що для абсолютної більшості представників іхтіофауни осіменіння і запліднення ікри відбувається у воді, поза материнським організмом. Така форма осіменіння і запліднення отримала назву зовнішнього, що знайшло відповідне відображення в спеціальній літературі. Проте деякі види риб характеризуються внутрішнім осіменінням і заплідненням. Цей процес проходить усередині материнського організму. При цьому в самців таких видів риб є спеціальні парувальні органи, які в найпростішому вигляді представлені анальною папілою. Самці у акул і скатів мають спеціальний апарат — птеригоподій, який сформувався із крайніх внутрішніх променів черевних плавців. Ці промені, збільшуючись в розмірах, утворюють на кожному із плавців масивний виріст, на внутрішньому боці якого є жолобок. При спарюванні самець складає обидва вирости разом і уводить їх в клоаку самки.
Серед костистих риб є достатня кількість живородящих видів, зокрема в ряду короподібних. Для цих видів риб характерна наявність видозмінених променів анального плавця (частіше третього і четвертого), які перетворені в досить складний копулятивний орган — гоноподій. У сома трахіхорієтеса відразу після внутрішнього запліднення відбувається викидання запліднених ікринок. Внутрішнє запліднення у деяких родів морських окунів також виключає внутрішньоутробний розвиток. Сперма цих риб зберігається в порожнині тіла самки до дозрівання яйцеклітин. Напевно, в даному випадку, внутрішнє запліднення сприяє заплідненню більшої маси ікри. Після запліднення, протягом короткого проміжку часу, проходить викид ікри в зовнішнє середовище, де відбувається подальший розвиток нащадків.
Таким чином очевидно, що сучасна іхтіофауна, сформована в процесі філогенезу, представлена досить великою видовою різноманітністю і характеризується величезною варіабельністю зовнішньої і внутрішньої будови, а також систем і органів певних видів, їх адаптацією до умов існування, що суттєво відбилось і на розмноженні риб.
Доцільно зробити висновок, що запліднення — це природний процес, але необхідною умовою запліднення і утворення зиготи є проникнення сперматозоїда в яйцеклітину, що можливо лише за безпосереднього контакту сперматозоїда і яйцеклітини. Це положення є загальним і не залежить від способу осіменіння. Воно правильне для зовнішнього і внутрішнього осіменіння, для натурального розмноження в природних умовах і для штучного розведення риб. Знання загальних закономірностей запліднення і його видоспецифічних особливостей стосовно окремих систематичних груп дозволяє не тільки усвідомлено підходити до оптимізації технології штучного розведення риб, але і різко поліпшувати умови відтворення цінних видів риб в природних і штучних водоймах, що виключно важливо для рибогосподарських водойм в умовах зростання антропогенного впливу.
Виходячи з викладеного, доцільно навести диференцировку сперматозоїдів залежно від існуючих критеріїв, основою яких є концентрація сперматозоїдів в 1 мм . Поряд з цим оцінюють тривалість руху сперміїв за наявною систематикою:
бал 5 - всі спермії рухливі, рух поступальний, рухливість дуже висока;
бал 4 - чітко виражені поступальні рухи, окремі спермії мають зигзагоподібні, коливальні рухи;
бал 3 - зигзагоподібні рухи переважають над поступальними, окремі спермії нерухомі;
бал 2 - поступальні рухи майже відсутні, є лише коливальні, окремі екземпляри демонструють коливні рухи, зрідка зустрічаються зигзагоподібні, до 75 % сперміїв нерухомі;
бал 1 - всі спермії нерухомі.
Оцінюючи сперму, звертають увагу на консистенцію, забарвлення, відсутність частин екскрементів, зайвої рідини, крові, що суттєво знижує термін зберігання та запліднюючу здатність.
Сперма різних видів риб зберігає рухливість, а з цього здатність запліднювати, протягом різного терміну. При правильному зберіганні сперми, вона не втрачає здатності запліднювати декілька днів.
Зрозуміло, що вибірковість запліднення існує, вона залежить від якісних характеристик сперматозоїдів. При цьому з урахуванням здатності ікри до запліднення необхідно акцентувати увагу на всіх особливостях батьків. Особини, які вперше досягли статевої стиглості, переважно демонструють сперму не дуже високої якості, що не виключає невисокої якості сперми для плідників старших вікових груп. Узагальнюючи це положення, необхідно акцентувати увагу на тому, що висока якість сперматозоїдів, за рівних факторів середовища і здатність до осіменіння яйцеклітин, типова для плідників середньовікових груп, на яких базуються штучні відтворювальні комплекси та нерестові популяції в природних умовах.
Одним з головних показників якості плідників є якість нащадків. Саме виходячи з цього постулату зрозуміло, що якість статевих продуктів, а безпосередньо сперматозоїдів, безумовно впливає на формування якісних і кількісних, екстер’єрних та інтер’єрних показників. При цьому необхідно враховувати і жіночу складову, яка в значній мірі корегує і може нівелювати вплив чоловічої статі.
Відповідно до тематики, доцільно привести народну говірку, яка безумовно належить фахівцям - «Від поганого семені не чекай доброго племені». Виходячи з цього ефективність запліднення, його безумовна вибірковість пов’язані з якістю сперматозоїдів, що орієнтує на сумнівність процесу в природних і трансформованих акваторіях і жорсткий штучний відбір з урахуванням віку статевозрілих особин при формуванні ремонтно - маточних стад відповідних рибничих господарств.
Розглядаючи проблему підбору плідників для уявлення і формування відповідного світогляду, необхідно пов’язати між собою фактично взаємопов’язані теоретично - технологічні заходи, а саме методи відбору та підбору.
Успіх селекції залежить від правильності оцінки риб при відборі їх для відтворення. Потомство відібраної риби повинно повторити показники батьків, чи переважати їх. Таким чином, відбір є рушійною силою поліпшення спадкових якостей тварин. Ефективність відбору визначається величиною мінливості, спадковості та інтенсивності відбору. За малої мінливості селекціонер може просто не знайти у стаді особин, які б відповідали його вимогам. Дуже значна мінливість також небажана, оскільки, проявляючись у кожному наступному поколінні, зумовлює підвищену величину регресії, тобто повернення до середніх показників популяції у нащадків тварин, які були попередньо відібрані за тією чи іншою бажаною ознакою. Чим вища спадковість, тим ймовірніший прогноз генотипу племінної риби при відборі.
В залежності від способу оцінки відібраних особин розрізняють два основних методи відбору: масовий та індивідуальний.
Масовий відбір - є основним методом селекції риб. При цьому оцінку і підбір здійснюють за масою тіла, екстер'єром та іншими ознаками, тобто їх фенотипом, передбачаючи, що «добрі» фенотипи мають і «добрі» генотипи. На плем'я залишають особин, які найповніше задовольняють бажаний тип, а інших вибраковують. Ознаки при відборі можуть бути найрізноманітнішими, а їх вибір залежить від мети селекції.
Основна перевага масового відбору полягає у його відносній простоті, що робить його доступним не тільки для фахівців, але і для досвідчених практиків. Селекціонер працює з чисельним матеріалом, що дає змогу досягати високої, достатньої ефективності. Проте оцінка за фенотипом за масового підбору має певні вади, вона не дозволяє достовірно робити висновок про генетичну цінність відібраної особини. Це можна здійснити лише за індивідуального відбору.
Індивідуальний відбір. Ґрунтується на оцінці фенотипу найближчих родичів. Опосередковане значення фенотипу родичів відібраної особини дає можливість визначити її генетичну цінність і тому відбір індивідуальний називають за генотипом. У селекції тварин, де досягнуті найвищі показники, використовують три типи індивідуального відбору: відбір за походженням, сімейний відбір і відбір за якістю нащадків.
При відборі за походженням враховують продуктивність родичів, що потребує систематичних записів, родоводу риб. За сімейної селекції нащадки від різних пар чи невеликих груп плідників вирощують в максимально ідентичних умовах. Потім визначають якість цих сімейств і вибирають кращі з них для подальшого вирощування та розведення. Оцінюють сімейства за середніми величинами, розрахованими для кожної сім'ї. Завдання сімейної селекції та індивідуального відбору дещо різняться між собою, оскільки у першому випадку кращі сім'ї, а в другому - кращих плідників. Вважається за бажане об'єднання цих двох форм відбору в єдиний процес, що значно підвищує швидкість досягнення мети і загальний ефект.
Не треба протиставляти одне одному - застосування індивідуального відбору не знижує ролі масового відбору. Виходячи з відомих наукових концепцій, доцільно впровадити комбінований відбір, який полягає у послідовному проведенні в одному поколінні сімейної селекції, масового та індивідуального відборів.
Відбір за нащадками. Найефективніший метод індивідуального відбору, який дозволяє отримати об'єктивну характеристику відносно якості плідників. У даному випадку оцінюваних плідників (самицю чи самця) парують із кількома плідниками іншої статі і за продуктивністю нащадків роблять висновок про племінну цінність плідника.
Результати дослідів щодо оцінки плідників залежать від фізіологічного стану риб, маси тіла, вгодованості. Більш крупні й вгодовані плідники дають кращих нащадків. При цьому батьківський і материнський ефекти особливо сильно проявляються у нащадків на ранніх стадіях розвитку: у коропа вплив самців проявляється в основному до досягнення нащадками віку 1-2 міс, вплив самиць - до кінця першого року вирощування. Фенотипове значення ознаки, за якою роблять висновок про племінну цінність тварин, залежить певною мірою від поєднання спадкових факторів та умов середовища. Взаємодія «генотип-середовище» особливо сильно виражена в ознаках із низькою спадковістю, зокрема ріст і виживаність. У риб особливо сильний вплив на результати оцінки відносної цінності різних груп може мати, наприклад, різна щільність посадки при вирощуванні.
При цьому виключне значення має низка умов, або технологічні параметри фону, на якому відбувається селекція. Технологія виробництва при селекції риб повинна бути подібною до технології потенційного товарного рибництва. Інакше у племінних господарствах формуватимуться типи риб, не придатних для товарних господарств, що, на великий жаль, має місце, нівелюючи зусилля селекціонерів.
Відбір за однією й тією ж ознакою, але у різних умовах, сприяє створенню особин, які суттєво різняться між собою за своїми спадковими якостями.
Зазначені вимоги до умов вирощування селекціонованого матеріалу поширюються лише на період, що передує основному відбору, наприклад, при селекції коропа за масою тіла - до досягнення рибами дворічного віку. В подальшому основним завданням стає вирощування фізіологічно повноцінних плідників, що досягається за рахунок оптимізації умов; створення оптимального фізико-хімічного режиму води, розрідженої посадки, годівлі високоякісними комбікормами, на тлі добре розвиненої природної кормової бази. Вирощені в таких умовах плідники можуть повною мірою проявити свої спадкові відмінності за репродуктивними властивостями (швидкість, статевого дозрівання, плодючість), що дає змогу вести відбір і за цими важливими ознаками.
Суттєвим моментом при організації масового відбору і оцінки плідників за якістю нащадків є стандартизація умов середовища; щільність посадки, годівля, тривалість вирощування. Припустимо як спільне, так і роздільне вирощування риб, при цьому необхідна не менш як триразова повторність дослідів. За сумісного вирощування риб різних ліній, сімейств і груп необхідно вирівняти посадкову середню масу. Якщо це неможливо, треба визначити поправочний коефіцієнт і внести відповідні нормативи в одержані прирости. Піддослідні групи риб повинні бути поміченими, таким чином, який гарантує їх від перемішування протягом дослідного періоду.
В основі всіх існуючих форм відбору покладено принцип використання генетичної мінливості. Ефект відбору за полігенними ознаками визначається і основними показниками: спадковістю ознаки, за якою ведеться відбір, і селекційним диференціалом.
Ефективність селекції визначається значною мірою застосуванням раціональних систем вирощування ремонтного молодняка. Система вирощування молоді риб повинна забезпечити нормальний ріст і розвиток організму, сприяти достатньо повній реалізації генотипу.
Методи підбору. Мета підбору полягає у складанні батьківських пар для одержання нащадків з бажаними властивостями. Підбір завершує всю попередню роботу по вирощуванню, виявленню господарської і племінної цінності підбору кращих особин для їх розмноження. Підбір - це синтез, у результаті якого селекціонер намагається найдоцільніше поєднати у нащадків основні бажані ознаки самців і самиць, відібраних для відтворення.
Підбір плідників заснований на відмінностях у ступені вираженості у спарюваних особин бажаних якостей. Парування самців і самок, які різняться між собою за ступенем вираженості ознаки, одержало назву різнорідного (гетерогенного) підбору. Основна мета його визначається концепцією про те, що гірше з кращим дає краще. Підбір може бутигетерогенним або гомогенним за віком, екстер'єром, екологічними умовами, в яких вирощувались спаровані особини. Всі ці фактори слід враховувати при підборі, але головними, що визначають доцільність підбору, є показники продуктивності плідників і можливість поліпшення їх при даному поєднанні.
Однорідний (гомогенний) підбір полягає в тому, що самці і самки при підборі подібні між собою за ступенем вираженості даної ознаки. Цей прийом дає змогу надійно відтворити в потомстві ознаки породи, тип та індивідуальні продуктивні якості батьків, зумовлює підвищення успадкування ознак.
Залежно від диференційованості підбору і від кількості спарованих самок і самців розрізняються підбір індивідуальний та груповий.
Індивідуальний підбір. Застосовують у спеціалізованих, племінних господарствах, де добре поставлений облік індивідуальних якостей плідників, його здійснюють фахівці високої кваліфікації. При його використанні слід визначити, яку з самиць і з яким самцем доцільно парувати, щоб в очікуваному потомстві одержати нові якості.
Груповий підбір. Одержав значне розповсюдження, його все ширше використовують у тваринництві, в тому числі і в рибництві. У спеціалізованих племінних господарствахосновним методом удосконалення тварин стає робота з лініями і сімействами, іншими спорідненими групами.
Лінія ведеться за самцями, оскільки максимальний відбір, особливо у малоплідних тварин, можливий лише серед самців, без збитків для чисельності тварин, яких розводять. Найважливіше завдання селекціонера, який працює з лінією, полягає у збереженні цінних і рідкісних генетичних комбінацій родоначальника лінії.
Мета розведення за лініями - розвиток і закріплення у нащадків цінних особливостей кращих тварин для одержання наступного покоління із стійкою спадковістю, використання якого забезпечить швидше удосконалення стада чи породи. Зробити це можливо не лише жорстким
відбором особин більш подібних до родоначальника, а й помірним інбридінгом. Застосування інбридінгу дає можливість використовувати одержану цінну генетичну комбінацію для створення більш чи менш однорідних груп. Тому при підборі необхідно враховувати притаманні цим групам властивості. Знання спадкових особливостей спорідненої групи є більш надійною основою для підбору і прогнозу його результатів.
Слід мати також на увазі, що в процесі індивідуального розвитку спостерігаються закономірні зміни морфологічних, фізіологічних, біохімічних та інших особливостей організму, причому час і порядок прояву цих змін в онтогенезі визначаються спадковістю організму. При одновіковому і різновіковому підборі плідників коропа якість потомства і вихід товарної продукції залежать і від віку риби. Використання у відтворенні крайніх вікових груп (тих, які нерестяться вперше і старших) приводить до одержання потомків з пониженою життєздатністю.
Поряд з викладеним достатньо цікавим в теоретичному і практичному плані є біологічне значення різноякісності стад риб. Ця особливість виступає в якості основи підбору плідників та технологічних складових, що суттєво у практичному рибництві.
Формування високопродуктивних маточних стад передбачає обов’язкову наявність в їхньому складі високого рівня гетерогенності, яка є біологічною основою удосконалення та прогресу на шляху домістефікації, формування порідних груп та порід. З метою забезпечення цієї умови в процесі селекційно - племінної роботи, фахівець постійно працюючи над забезпеченням оптимального співвідношення статевих та вікових груп, приділяє прискіпливу увагу питанням гетерогенності за відповідними ознаками, виходячи із бажаності її забезпечення і накопичення.
Виконуючи технологічні операції в процесі осіменіння ікри, підбирають самиць і самців, користуючись відповідними параметрами гетерогенності. В практичному плані реалізація сформованої концепції зводиться до того, що для отримання високоякісної молоді, здатної
демонструвати високу життєстійкість та продуктивні якості, необхідно на всіх технологічних операціях, починаючи з відтворення, і, завершуючи наступною генерацією плідників, орієнтуватися на створення стада плідників з різноякісними позитивними особливостями, які кореспондуються з якісними ознаками, забезпечуючи відповідну спадковість.
Створення і тривала робота, яка використовувалася протягом багатьох років, була орієнтована на оптимальну якісну структуру самиць і самців у стаді плідників і базувалася на вимозі так званого «прилиття крові». При цьому мається на увазі залучення аналогічних плідників з інших господарств, інших регіонів, виловлених з природних акваторій.
В практичному рибництві господарства обмінюються плідниками, але зручніше обмінюватися спермою відповідних плідників, або заплідненою ікрою, що не вимагає суттєвих витрат на транспортування плідників і пов’язані з цим травматизація та відхід особин в процесі транспортування.
З попередніх спеціальних дисциплін, в першу чергу з курсу «Розведення риб» загальновідомо, що еколого - фізіологічні аспекти біотехніки штучного і природного відтворення базуються на однаковій нейро - гуморальній основі. Поряд з цим ефект досягається в природних умовах без втручання людини, а в штучних умовах на тлі відсутності відповідних екологічних передумов, людина втручається в цей процес активно, але втручання має природний, за своїм походженням і дією характер.
Запропоноване теоретичне обґрунтування має принциповий характер, фактично є підґрунтям сучасних уявлень про процеси, які пов’язані з відтворенням в природних, трансформованих і штучних умовах.
Виходячи з викладеного, бажаним є короткий огляд еколого- фізіологічних аспектів біотехніки штучного і природного відтворення риб. Ефективність відтворення, як і сам факт відтворення можуть бути різними, відтворення взагалі може не відбутися, що у свою чергу залежить від багатьох факторів зовнішнього середовища з якими тісно пов’язані фізіолого - біохімічні процеси плідників і особливо чітко простежуються у пойкілотермних тварин, до яких належать риби.
Штучне відтворення або розведення риб передбачає застосування відповідної біотехніки, яка у свою чергу базується на проведенні низки послідовних технологічних операцій. При цьому розрізняють загальну технологію, яка не несе в собі видоспецифічних особливостей відповідних видів риб, і спеціальна технологія штучного відтворення конкретних видів риб.
Еколого - фізіологічні аспекти біотехніки розведення і розмноження риб передбачають наявність відповідних факторів середовища з одного боку і наявності статевозрілих особин з другого боку, що орієнтує на певні положення.
Зрозуміло, що в природних умовах коли відсутність негативного впливу людини забезпечується збереженням біологічної норми, розмноження відбувається на загальному позитивному фоні. Відносно природної динаміки факторів середовища, то в переважній більшості випадків особини видів адаптовані до таких явищ в процесі філогенезу.
В штучних умовах, стикаючись з проблемою розведення, виникає необхідність штучно формувати, в припустимих межах, екологічних параметрів середовища, що дозволяє успішно працювати з видами, які невибагливі до умов нересту і переважно є еврибіонтами. Працюючи з видами риб, які віднесені до стенобіонтів, управління екологічними параметрами середовища, в межах можливостей, не забезпечує досягнення ефективного розмноження, що спонукає до використання фізіологічно активних препаратів відповідного спрямування.
Сприйняття попередньої інформації дозволяє безпосередньо перейти до загальних технологічних процесів, які пов’язані з розведенням риб.
- Наявність статевозрілих самців і самиць, які можуть бути безпосередньо в межах підприємства або заготовлюватися в природних (трансформованих) акваторіях.
- Переднерестове утримання плідників в умовах максимально наближених до природних.
- При досягненні переднерестових температур для конкретного виду висадити нерест, попередньо забезпечивши наявність відповідного субстрату. Види, які демонструють певну вибагливість до умов нересту, їх забезпечення не є реальним в штучних умовах, доцільно використовувати фізіологічно активні препарати гонадотропної дії.
- Керуючись видоспецифічними особливостями, динамікою температури води, через певний період після ін'єктування отримують статеві продукти.
- Залежно від особливостей виду здійснюють осіменіння одним з існуючих способів і створюють умови для ефективного запліднення.
- Залежно від особливостей виду, типу інкубаційних апаратів, необхідності запліднення ікри її завантажують в інкубаційні апарати, в яких відбувається ембріогенез.
- Збір і витримування передличинок або вільних ембріонів до переходу на личинкову стадію розвитку.
- Пересадка личинок у штучні і природні акваторії, що залежить від цільового призначення.
За наявності всіх рівних факторів в умовах природного і штучного нересту його ефективність і якісні характеристики потомства безумовно залежать від дотримання біотехніки штучного відтворення і умов природного нересту, умов попереднього нагулу плідників, якісних характеристик плідників, їх статевих продуктів. На цьому фоні практика зіткається з показниками, які суттєво відхиляються від нормативних за наступних параметрів: запліднюваність ікри, вихід вільних ембріонів, передумовою якого є концентрація сперматозоїдів, термін їх контакту з ікрою, не може бути виключений і людський фактор.
Для плідників, які приймають участь у нересті, особливо для видів з весняно - літнім нерестом, виключного значення набуває вгодованість і жирність. Ця вимога базується на тому, що в переважній більшості випадків в період зимівлі плідники не живляться і активно втрачають запаси жиру в зимовий період. За відсутності відповідних жирових запасів, які накопичуються влітку, в період нагулу, плідники виснажуються, хворіють і не здатні продукувати високоякісні яйцеклітини і сперматозоїди, що орієнтує на створення відповідних умов для плідників і період інтенсивного нагулу.
Аналіз процесу еколого - фізіологічних основ природного і штучного відтворення риб переконливо свідчить про те, що поряд з об’єктивними передумовами біологічного характеру важливою складовою є оператор-фахівець, від якого залежить безпосередньо виконання технологічних операцій.
Статеві продукти високої якості можуть бути зіпсовані недбалим відношенням при їх одержані, зберіганні, використанні. Саме інє'ктування вимагає певної кваліфікації, яке виключає травмування риби і одночасно виключає витікання препарату у зовнішнє середовище після ін’єкції.
Особливу увагу необхідно приділяти безпосередньо роботі з плідниками, використовуючи для цього відповідний інвентар і мати відповідні навички.
Плідник - продукт тривалої людської праці протягом багатьох років, його збереження та участь в багатьох нерестових кампаніях свідчить про професіоналізм фахівця, який з ним працює, свідчення свідомого дотримання біотехніки штучного розведення риб не тільки за формальними ознаками, а одночасно з турботою про перспективу.