ТЕОРЕТИЧНІ ОСНОВИ РИБНИЦТВА - І.М. Шерман - 2011
2.ТЕОРЕТИЧНІ ОСНОВИ ФОРМУВАННЯ ТА ВИКОРИСТАННЯ ПРОДУКТИВНИХ ВЛАСТИВОСТЕЙ РИБ
2.3.Закономірності формування статі та статевих продуктів риб
Висока сучасна чисельність видів риб сформувалася протягом тривалого філогенезу і напрацювала ряд особливостей в межах ареалу. Виходячи з цього, певний теоретичний інтерес викликає система понять про внутрішньовидові біологічні угруповання риб, що є вагомою складовою формування статі та статевих продуктів риб.
Час нересту у різних видів риб досить різноманітний і приурочений до різних сезонів року. Промислових риб помірних та високих широт розділяють на дві групи, нерест яких відбувається у різні пори року.
Перша група риб нерестує у березні - липні, а друга - у вересні - січні. У обох груп риб викльовування передличинок приурочені до весни і літа, коли у водоймах відбувається інтенсивний розвиток кормового планктону, який є їжею для всіх видів риб на ранніх стадіях їх розвитку. Неоднакові календарні терміни ікрометання властиві також рибам одного виду і навіть однієї популяції.
Прохідні риби йдуть на нерест в ріки в різний період року і з неоднаковою ступінню стиглості статевих залоз. Одні з цих риб нерестять невдовзі після заходу в річку, інші - лише на наступний рік. Спостерігаючи таке явище, Л.С.Бер дійшов висновку, що у деяких лососевих і осетрових риб існують ярові і озимі раси. Ярові риби мігрують із моря в ріки, де вони мечуть ікру в цьому ж році. Озимі риби йдуть із моря в ріки, де вони нерестять лише наступного року.
Вивчивши особливості розмноження куринського і волзького осетра, вченими Н.А. Гербильским, Б.Н. Казанским, И.А. Баранниковою встановлена наявність біологічних груп всередині їх популяції, що слугувало поглибленню вчення Л.С. Берга про сезонні раси. Було встановлено, що осетер, мігруючий із Каспію у Волгу, має чотири біологічних групи: ранній яровий осетер, пізній яровий осетер, озимий осетер літнього ходу і озимий осетер осіннього ходу.
Ранній яровий осетер входить у Волгу ранньою весною з гонадами в близькій до завершення або завершеної IV стадії стиглості і відкладає ікру невдовзі після підйому в річку, коли температури досягають оптимальних для нересту значень, а це середина травня - початок червня. Пізній яровий осетер входить у Волгу весною і влітку з гонадами у незавершеній IV стадії стиглості і нерестить у цьому ж вегетаційному сезоні після зниження високих літніх температур, а саме в липні - серпні.
Озимий осетер літнього ходу мігрує до Волги в травні - липні зі слаборозвиненими статевими продуктами, яєчники знаходяться в III - IV, а сім’яники в II - III стадіях стиглості і відкладає ікру ранньої весни наступного року, що припадає на кінець квітня - травень. Озимий осетер осіннього ходу мігрує до Волги в серпні - вересні з гонадами в близькій до завершення IV стадії зрілості і нерестить так же, як і озимий осетер літнього ходу, ранньою весною наступного року, що відбувається в кінці квітня - в травні.
Всі вказані екотипи мігрантів, за виключенням пізнього ярового осетра, є російським осетром. Пізній яровий осетер представлений в основному особинами другого осетра - перським осетром. Крім куринського і волжського осетра, внутрішньопопуляційна диференціація існує у осетра і в ряді інших рік. Біологічні групи є також всередині популяцій севрюги і білуги в ріках Каспійського і Азовського басейнів.
Озимі форми осетрових з яєчниками в IV стадії стиглості, в яких ооцити досягли дефінітивних розмірів, а це кінець трофологічного росту, заходять в річку і там в ній зимують. Ооцити цих риб знаходяться у такому стані аж до весни. З настанням сприятливих екологічних умов для нересту цих риб відбувається завершення оогенезу під впливом гонадотропного гормону гіпофізу. Такого роду адаптація, яка здійснюється за рахунок затримки ооцитів в стані завершеного протоплазматичного росту, залежить від функції клітин мезоаденогіпофізу, які продукують гонадотропний гормон. Восени вміст цього гормону в гіпофізі озимих форм осетрових у два рази нижче, ніж весною і влітку у ярових.
Велике значення в адаптації певної популяції деяких видів лососевих риб, а саме сьомги, до умов розмноження має утворення карликових самців в її межах, які постійно знаходяться в річці і приймають участь у нересті з мігруючими самками.
Місця нересту озимих риб розташовані у верхній течії рік, а ярових риб - у нижньому. Така внутрішньовидова диференціація прохідних риб відносно місць нересту також носить пристосувальний характер і дозволяє використовувати нерестовища, розташовані у різних ділянках річки на дуже великій її протяжності, що забезпечує сприятливі умови для виживання нащадків. Після нересту плідники прохідних риб і їх молодь мігрують із рік до моря.
Наявність біологічних груп всередині популяції обумовлено умовами ареалу. Внутрішньопопуляційна диференціація риб не проявляється за відсутності необхідних умов для її розвитку.
Таким чином, диференціація у межах популяції - це пристосування риб до умов розмноження і розвитку. Внутрішньовидова диференціація - це найважливіша видова адаптація до умов ареалу, що приводить до найбільш повного використання її можливостей.
Знання внутрішньопопуляційної диференціації дозволяє не лише науково обґрунтовано вирішити питання щодо регулювання рибальства і лімітування вилову цінних прохідних промислових риб, але й правильно розташовувати рибничі підприємства та визначати їх потужність. Це дає можливість складати сезонний графік роботи з розведення різних біологічних груп в необхідній кількості і співвідношенні з тим, щоб зберегти генетичну і екологічну структуру популяцій і збільшити їх чисельність.
Збереження внутрішньопопуляційної диференціації у риб особливо важливо в сучасних змінних умовах під впливом негативних природних і антропогенних чинників, коли спостерігається скорочення нерестових площ і співпадання місць нересту у багатьох біологічних груп риб, порушення динаміки ходу плідників в ріки і погіршення їх фізіологічного стану. У зв’язку з цим необхідна подальша інтенсифікація риборозведення, яка передбачає впровадження у виробництво нових прийомів і методів управління біологічним процесом.
Знання внутрішньопопуляційних зв’язків у риб дозволили здійснити і науково обґрунтувати та розробити метод регулювання ходу статевого циклу і вистигання плідників осетрових, що дозволило також здійснювати розведення всіх біологічних груп цих видів риб у промислових масштабах. Впровадження цього методу у виробництво дозволяє раціонально управляти осетровим господарством у південних морях, має вагомі значення в процесі акліматизації та реакліматизації.
Штучне відтворення риб фактично є технологією, складові якої передбачають управління статевим циклом риб.
При промисловому риборозведенні велике значення має логічне планування і організація роботи з підтримання і збільшення чисельності
тієї чи іншої популяції з урахуванням її структури, яка історично склалась, що є важливою пристосувальною властивістю виду у кращому використанні нагульного і нерестової частин ареалу. У зв’язку з цим при промисловому риборозведенні слід зберігати всередині кожної популяції риб певне співвідношення між різними біологічними групами, які необхідні для прогресу виду і бажані для людини.
Завдяки проведеним досліджень багатьма вченими ця проблема успішно вирішується в лососівництві та осетрівництві. Протягом останніх років були розроблені теоретично і впроваджені у виробництво еколого - фізіологічні основи управління статевим циклом осетрових. Вже зараз рибничі заводи можуть управляти вистиганням плідників різних біологічних груп осетрових. Виходячи з цього, напрацьовані відповідні варіанти витримування різних біологічних груп осетра у спеціально побудованих з цією метою басейнах на осетрових рибничих заводах.
- Витримування протягом декількох діб плідників раннього ярового осетра, гонади яких знаходяться в IV стадії стиглості, в басейнах з певним водообміном і за природних температур річкової води без зміщення їх статевого циклу.
- Витримування протягом 1,5-2 міс. плідників пізнього ярового осетра і протягом 7-8 міс. плідників озимих форм осетра до переднерестового стану, а саме IV стадія стиглості гонад) в басейнах з певним водообміном і за природних температур річкової води без зміщення їх статевого циклу.
- За оптимальних нерестових температур в річці можна здійснювати резервування до 2-3 міс. плідників ярового осетра, гонади якого знаходяться в IV стадії стиглості, і попередньо витриманих на заводі протягом 7-8 міс. плідників озимого осетра літнього і осіннього ходу, у котрих яєчники і сім’яники також мають IV стадію стиглості. Утримання цих плідників в басейнах з регулюємою температурою води, поступово зниженою до 2-4о С, і певним водообміном дозволяє змістити статевий цикл на більш пізні терміни шляхом його
затримки на IV стадії стиглості гонад. За потреб рибничого підприємства в цих плідниках температуру води в басейнах поступово підвищують до нерестової. Отже, за витримування плідників осетра в басейнах за регулюємих температур води і певного водообміну можна затримати розвиток гонад в IV стадії стиглості, за якої риби досягають переднерестового стану. Це забезпечує зсув у статевому циклі плідників осетра на бажаний час.
У всіх варіантах кінцеве вистигання статевих продуктів плідників різних біологічних груп осетра, гонади яких знаходяться в IV стадії стиглості, відбувається, як і у інших видів осетра, протягом 1-2 діб шляхом введення їм гонадотропного гормону гіпофіза, або препаратів аналогічної дії.
Таким чином, на думку А.П. Іванова, рибоводи можуть отримати зрілих плідників різних біологічних груп осетра, застосовуючи той чи інший варіант управління їх статевим циклом.
Поряд з еколого - технологічними складовими, фізіолого - біохімічними чинниками важливе значення у формуванні сучасних технологій мають генетичні основи формування статі риб.
Як і всі видові особливості, статева структура популяції і її зміни у зв’язку із змінами умов життєдіяльності організму, зв’язані не лише з хромосомним набором, але й являє собою пристосувальну реакцію до умов оточуючого середовища. Раніше існувала точка зору про те, що співвідношення статей змінюється лише в результаті природного відбору без урахування впливу на ці процеси оточуючого середовища. Багаточисельними дослідженнями було встановлено, що шляхом зміни температури, солоності води, щільності посадки риб, зміни інтенсивності і характеру обміну речовин, а також впливом на ікру, що розвивається, статевими гормонами, можна змінювати співвідношення статей в популяції. Користуючись цими принципами, формування статей в популяції рибоводи можуть на свій розсуд, в залежності від завдань регулювати статевий склад вирощуваного стада риб.
Планування відповідних складових технологічного процесу передбачає отримання знань щодо закономірностей динаміки плодючості риб і зміни, які розглядаються як пристосувальна реакція популяції на зміну забезпеченості риби їжею. Адже відомо, що за покращення забезпеченості риби їжею плодючість популяції зростає, а за зменшення, навпаки, скорочується. Виходячи з цього, вкрай важливо розуміти закономірності розвитку статевих продуктів риб, оскільки від цього залежить успіх отримання повноцінних ікри і молок.
Розвиток статевої залози - це процес, який відбувається не сам по собі, а у єдності зовнішнього і внутрішнього, який залежить як від попереднього стану, так і від тих умов, в яких знаходиться організм в процесі вистигання статевих залоз. Розвиток останніх, як відомо, нерозривно зв’язаний із загальним обміном речовин. Враховуючи це, шляхом регуляції обміну речовин можливо в певних межах регулювати хід розвитку статевих залоз.
Необхідно також враховувати і ті обставини, що гаметогенез у різних видів риб достатньо специфічний, що необхідно враховувати поряд з тими умовами, які необхідні для проходження кожної фази овогенезу і сперматогенезу.
Успішність проведення технологічних заходів передбачає володіння відповідними знаннями, які розкривають закономірності зміни якості статевих продуктів. Як і плодючість, якість статевих продуктів виробляється як пристосування до умов життя і змінюється зі зміною цих умов. Так у плітки, за певного режиму живлення, найбільший запас жовтка в ікрі спостерігається у самок у віці п’яти років. За зміни режиму живлення, можна отримати ікру з найбільшим запасом жовтка, як у самок старшого віку, так і в більш молодих риб. В ікрі більш жирних риб, які, зазвичай, є більш плодючими, як правило, міститься дещо менший запас жовтка. З викладеного логічно витікає, що регулюючи режим нагулу, можна регулювати як вік статевого визрівання риб, так і потенційну плодючість.
Відтворення риб в штучних умовах передбачає наявність розмірно - статевої структури популяції. Загальне співвідношення статей у популяції багатьох видів риб близько 1:1, але співвідношення статей у різних вікових і розмірних вікових групах виявляється досить різним. Переважання самців спостерігається у молодших вікових групах, що пов’язано з більш раннім їх статевим достиганням порівняно з самицями, а також більш раннім вступом до складу нерестового стада і меншою тривалістю життя. Камбали Північного моря у своєму складі мають кількість самців у дволітньому віці - 63%, а потім їх кількість поступово зменшується, складаючи у 5-6-річному віці 50-51%, а в 9-річному - менше 13%. Розглядаючи певні нерестові популяції, можливо констатувати, що таке явище, як переважання дрібних самців у нерестовій популяції властиво далеко не всім видам риб. У деяких видів сомових самки більш дрібні, ніж самці і їх відсоток вище серед дрібних риб у нерестовій популяції.
За характером співвідношення статей серед особин різних розмірів виділяють три групи. У представників першої групи, до яких віднесений океанічний оселедець, між самцями і самками не виявлено суттєвої різниці за темпом росту, віком статевого вистигання і тривалістю життя.
Другий тип зміни співвідношення статей з розмірами і віком характерний для осетрових, більшості коропових, багатьох лососевих, окуневих. У риб з цим типом співвідношення статей самці стають статевозрілими раніше самиць і, зазвичай, раніше помирають. У зв’язку з цим у них серед дрібних молодих статевозрілих риб переважають самці, а серед крупних - самиці. Переважання самиць серед крупних риб спостерігається і у риб в межах однієї вікової групи. Така структура популяції, коли самиці виявляються крупнішими самців, забезпечує більшу плодючість стада, оскільки крупні особини відкладають більше ікринок. У той же час завдяки тому, що самці дрібніші самиць, за однієї і тієї ж кормової бази забезпечується висока чисельність популяції за збереження її відтворювальної здатності. У більшості видів цієї групи за нересту з більш крупними самицями нерестять декілька більш дрібних самців.
Третя група риб - це види, у яких самці більш крупніші, ніж самиці, і вони переважають серед риб більш крупних розмірів. При цьому інколи виявляється, що за віком співвідношення статей виявляється оберненим. Більші розміри і більший вік самців порівняно з самками спостерігається у тих видів риб, у яких самці охороняють ікру, що характерно для сомів, губанів, бичків і деяких інших. У прохідних лососей більші розміри самців очевидно пов’язані з необхідністю подолання високих швидкостей течії протягом більш тривалого часу, що витікає з участю одного самця в нересті з декількома самицями.
У багатьох видів риб поряд з крупними за розмірами самцями, які інколи перевищують розміри самиць, є дрібні карликові форми самців про що сказано вище і типово для лососей. Такі самці у складі популяції в окремих випадках бувають представлені лише карликовими формами, що характерно для глибоководних вудильщиків.
У багатьох прохідних риб у мігруючій частині популяції відсоток самиць буває набагато вищим відсотку самців, що відмічено у багатьох лососей, гольців, аральського вусача. У деякі роки кількість самок досягає 80%. Виходячи з того, що в осіменінні ікри крупних самок приймають участь головним чином карликові самці, на рибничих заводах сперма карликових самців також може заміняти сперму прохідних самців, що технологічно має певний сенс.
У деяких випадках популяція може бути представлена двома групами риб обох статей - карликовими самцями і самицями і крупними прохідними самцями і самицями. Нерест крупних і карликових риб відбувається одночасно, але прохідні самці лосося після нересту, зазвичай, гинуть, а карликові нерестують декілька разів у житті.
У складі іхтіофауни переважають роздільностатеві види, але для багатьох видів характерним є гермафродитизм. Таке явище притаманне
багатьом костистим рибам. Серед гермафродитів є групи функціональних або синхронних і нефункціональних гермафродитів.
У гонадах функціональної групи гермафродитів можна розрізнити яєчникову і сім’яникову частини. В один і той же час в гонадах цих риб можуть знаходитись як зрілі яйця, так і сперма. Між тим у цих риб, зазвичай, самозапліднення не відбувається і кожна особина виконує почергово то функцію самки, то функцію самця. Вважають, що самозаплідненню в природних умовах заважають, очевидно, якісь не вивчені до цього часу фізіологічні бар’єри.
У риб, які належать до групи не функціональних гермафродитів, в гонаді також можна розрізнити дві частини: яєчникову і сім’яникову. Однак, на відміну від функціональних гермафродитів, у нефункціональних гермафродитів обидві частини не функціонують одночасно. У одних видів риб у молодому віці найбільшого розвитку досягає яєчникова частина, а самцева залишається неактивною і такі особини функціонують як самиці. Після одного або декількох ікрометань яєчник цих риб піддається редукції, овоцити резорбуються, а сім’яник досягає найвищого розвитку, а явище зміни статі носить назву протогінії.
Одночасно у складі іхтіофауни існують види, де спостерігається інша закономірність - у молодому віці вони функціонують як самці, а в старших вікових групах трансформуються у самиць. Така зміна статей носить назву протандрії.
У деяких видів риб (Diplodus annularis) є як роздільностатеві особини, так і гермафродити, причому процент гермафродитних особин з віком зменшується. Серед чотирирічних риб самиць 60%, самців 20%, гермафродитів 20%, а серед шестирічних кількість гермафродитів становить біля 3%.
Існує третя група риб - гермафродити, які у більш молодому віці мають яєчник і вони функціонують як самиці, а через деякий час у них спостерігається реверсія статі і старші вікові групи риб представлені лише самцями. У цих риб спостерігається також протогінія, однак, на відміну від протогінії нефункціональних гермафродитів, у риб з потенційним гермафродитизмом в гонаді самки не виражена сім’яникова частина. Серед овоцитів є лише недиференційовані статеві клітини - гонії, подальший розвиток яких і утворює замість яєчника сім’яник.
Біологічна суть гермафродитизму до кінця ще не встановлена. Між тим немає сумніву в адаптивному значенні цього явища. Протогінію і співвідношення статей, яке зсунуте в бік переважання самок у більш молодому віці плідників риб, можна розглядати як пристосування до збільшення швидкості відтворення за вкрай несприятливих умов. Так, у Monopterus albus, який вирощується на рисових чеках острова Ява, період розмноження досить короткий. Тому цей вид пристосувався до нестабільних умов на рисових полях. Після сухого періоду відбувається швидке відновлення популяції цих риб завдяки тому, що всі кормові ресурси використовуються в першу чергу для забезпечення процесів життєдіяльності особин, які функціонують як самиці, що і приводить до різкого підвищення чисельності популяції. Особини, які пережили несприятливий період, стають самцями, вони більш крупні, продукують більшу кількість сперми і здатні нерестувати з декількома самицями.
Отже, протогінія - це пристосування, яке забезпечує зростання чисельності популяції завдяки використанню кормових ресурсів водойми особинами, які функціонують як самки. Припускають, що протогінія притаманна тим видам риб, які живуть у водоймах або в біотопах з нестабільними екологічними умовами, або з нестабільною кормовою базою.
Значно рідше зустрічається явище протандрії. Припускають, що воно викликається дефіцитом кормової бази для вистигання самиць у більш молодому віці, але успішно використовується для визрівання самців. Але через деякий час самці, які підросли, перетворюються у крупних і, отже, у більш плодючих самиць.
Важливим позитивом явища гермафродитизму серед риб є й те, що у випадку синхронних гермафродитів забезпечується можливість відновлення популяції навіть за збереження поодиноких особин.
Щодо механізмів зміни статі у риб-гермафродитів, то він до цього часу досконало ще не вивчений. Існують лише припущення про те, що зміна статі відбувається під гормональним впливом. Адже доведено, що стать риб у значній мірі визначається статевими гормонами і будь-який агент чи вплив чинників водного середовища, які ведуть до зниження секреції гормонів, викликає перетворення статі. Реверсія статі може також спостерігатись у риб старших вікових груп у зв’язку із згасанням статевої функції, яке пов’язане з віковими змінами в обміні речовин.
Працюючи в обраній галузі, необхідно мати професійну точку зору на пристосувальне значення змін співвідношення статей і механізми регуляції статевої структури популяції.
Статева структура популяції має дуже важливе значення у відтворенні виду або популяції. У риб, як і в інших тварин, виробляються відповідні механізми регуляції статевого складу групи у зв’язку зі змінами умов життєдіяльності, зокрема, зі зміною забезпеченості організму їжею. Оскільки забезпеченість особини їжею тісно пов’язана з чисельністю популяції, то зміни чисельності повинні певною мірою відбиватися на співвідношенні статей в популяції, змінюючи її відтворну здатність. Існують літературні джерела про наявність тісного зв’язку між змінами статей і чисельністю популяції у круглоротих і риб. Зокрема встановлено, що у роки з більшою чисельністю популяції відсоток самців вище відсотку самок прісноводної форми морської міноги та деяких інших видів риб. У малочисельних нерестових групах горбуші самиці протягом багатьох років переважали над самцями, а в роки відносно високої чисельності, навпаки, самців було помітно більше, ніж самиць.
Основними причинами зміни співвідношення статей у риб є забезпеченість їх їжею і, очевидно, її якістю. За кращих умов відгодівлі співвідношення зсувається в бік самок, за гірших - в бік самців.
Збільшення кількості самців пов’язують з бідною кормовою базою, недостатньою для визрівання великої кількості самок.
Але у деяких видів риб зниження забезпеченості їжею викликає, навпаки, появу великої кількості дрібних самок, які рано визрівають, і здатні підтримувати високу чисельність навіть за несприятливих екологічних умов. Існує припущення про те, що у деяких випадках зміна співвідношення статей, зокрема у окуня, обумовлюється селекцією засобів лову. Однак, все ж-таки вважають, що основною причиною цих змін у окуня є різна забезпеченість їжею. За поганих умов живлення виділяються дві форми окуня, які швидко і повільно ростуть, і багаточисельні дрібні тугорослі самиці, які слугують їжею більш крупним самицям.
Існують також дані про те, що кількість самиць в популяції зростає за високих щільностей посадки риб. Виявляється, що і в потомстві риб, які живуть за високих щільностей посадки, самиці можуть за кількістю переважати кількість самців.
У процесі становлення водосховищ також спостерігається зміна співвідношення статей. В перший рік залиття Можайського водосховища суттєво покращилися умови відгодівлі риб, що викликало прискорення процесу розвитку статевих залоз риб. На другий рік залиття в нерестовий період популяції плітки кількість самців, які визрівають раніше самок, зросла більш ніж удвічі і досягла 59% замість 23,7%, як було раніше.
Інколи створюється ситуація, коли зі зміною щільності популяції змінюється темп статевого визрівання. При цьому, якщо зміни віку статевого визрівання в обох статей носять однаковий характер, тоді співвідношення статей у різних розмірних і вікових категоріях суттєво не змінюється. У випадку, коли темпи статевого формування статей у самців і самиць неоднакові, то це відбивається й на розмірно-статевій структурі популяції. Так, у круглого карася за виключно сприятливих умов росту різниця в часі визрівання самців і самиць нівелюється хоча за звичайних умов самці визрівають дещо раніше самиць, що не виключає одночасного статевого достигання. За цих умов відносна чисельність самців серед
молодших вікових груп зменшується, а серед старших, навпаки, - зростає. Співвідношення статей у нерестуючої частини стада може суттєво змінюватися в результаті пропуску самицями нересту за несприятливих екологічних умов чи умов нагулу.
Отже, на зміну забезпеченості їжею популяція реагує зміною співвідношення статей і розмірно-статевої структури популяції. Ці зміни можуть відбуватися в результаті як трансформації статі у риб, так і різного у часі статевого визрівання самців і самок або пропуску нерестового сезону, зазвичай самицями, за погіршення забезпеченості їжею.
В практичній діяльності існує певний інтерес до співвідношення самиць та самців в природних популяціях і в умовах культивування певних видів риб, що орієнтує на доцільність відносно теоретичної складової способів регуляції статі у риб.
Ученими та практиками у сфері рибництва нагромаджено значний досвід щодо механізмів та принципів регуляції статевої структури популяції у риб. Зміна співвідношення статей у популяції відповідно із змінами забезпеченості їжею безперечно вказує на те, що співвідношення регулюється посередництвом зміни перебігу обміну речовин. Основні механізми статі можна відобразити наступною схемою:
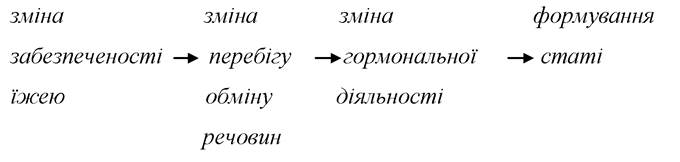
Вважають, що це є основним механізмом регуляції статевої структури популяції. Очевидно, що суттєве значення має зміна віку статевого вистигання і максимального віку у зв’язку зі зміною забезпеченості їжею.
З викладеного витікає думка відносно визначення характеру і становлення розвитку тієї чи іншої статі. З цього питання існує декілька точок зору. Так, одні дослідники вважають, що провідна роль належить статевим хромосомам або статевому хроматину, який був знайдений у
деяких тварин. Проте, статеві хромосоми вірогідно відомі лише у двох видів риб. Генетичне визначення статі у деяких видів риб автори хромосомної теорії намагаються довести дослідженнями з гібридизації.
Інші дослідники, виходячи з позиції єдності організму і середовища, розглядають статеву структуру і її можливі зміни як пристосувальний процес, який регулюється через обмін речовин і гормональну діяльність організму. У визначенні статі безперечно певну роль відіграє хромосомний комплекс, але не ізольовано, а в системі загального обміну. Отже, стать організму риб формується в процесі розвитку і в результаті взаємодії з середовищем, в результаті перебудови обміну речовин і регуляції гормональної активності.
Значна кількість досліджень присвячена проблемам регуляції статі в потомстві у риб з застосуванням гормональних препаратів. Особливої уваги заслуговують дослідження японського дослідника Ямамото, яким виконані роботи на рисовій рибці медаці. Він додавав до корму личинкам рисової рибки метилтестостерон, наслідком чого відбувалося перетворення самок у самців. Додаючи до корму естрон або стілбестрол, вчений викликав перетворення самців у самок. Від перетворених риб ним були отримані нащадки.
Введенням у воду тестостерону або естрогену, де знаходилася тільки що запліднена ікра або у воду, де відбувалося запліднення ікри, американський дослідник Барроу отримував в одному випадку самців, а в іншому - самок чавичі.
Деякими дослідниками з метою регуляції статі статевозрілим рибам, які були представлені мечоносцями, підшкірно вводили препарати еругону і тестостерону, наслідком чого було перетворення статевозрілих самок у плодючих самців. Живлення риб естрогеном також приводило до зміни співвідношення статей в бік зростання чисельності самок. Дослідженнями також встановлено, що у самців жіночий статевий гормон або викликає утворення змішаної статевої гонади (testis-ova), або гальмує розвиток сім’яника, або подовжує ювенільну жіночу стадію. Статевий гормон, який представлено андрогенними речовинами, може прискорити розвиток сім’яника у самців, але у самиць він викликає або перетворення статі, або гальмує розвиток, або зовсім не викликає впливу.
У цілому гормони суттєво змінюють збалансованість обміну речовин в організмі риб. Проте, ефективність біологічної дії гормонів визначається дозою препаратів. Помірні дози жіночого статевого гормону викликають перетворення самців гуппі у самиць, а високі дози приводять до кастрації. Оскільки забезпеченість їжею залежить не лише від кількості корму, але й від умов його споживання і засвоєння, то через зміну температури, яка впливає на перебіг обміну речовин, вдається впливати на співвідношення статей у риб.
Суттєвий вплив на процеси розвитку і росту риб має температурний чинник. Відомо, що розмноження риб відбувається в певній зоні коливань температури. За оптимальних температур для даного виду риб спостерігається нормальний розвиток ікри. Більш низька, порівняно з оптимальною, температура викликає затримку розвитку і вилуплення ембріонів із оболонок, а більш висока температура приводить до прискореного розвитку і більш раннього викльовування. Шляхом зміни температури вдалося здійснювати регуляцію статі риб. Впливом високої температури на двомісячних мальків Xiphophorns helleri вдалося викликати переважання серед отриманої молоді самців, а співвідношення статей було 5:1. Цей факт вчений Теріан пояснює руйнівною дією підвищеної температури на яєчник і перебудовою його в сім’яник. Інші дослідники, вивчаючи вплив температури та деяких інших чинників водного середовища на співвідношення статей, відмічають загальну тенденцію зростання чисельності самиць за інкубації ікри за більш високої температури води.
Певний вплив на співвідношення статей має солоність води. Ліндсей у трьохголкової колюшки за розвитку молоді у прісній воді отримував співвідношення статей 1 : 1, а в прісній + морській воді у співвідношенні 1 : 1 отримано 7 самок і 3 самців, у прісній воді + морській воді у співвідношенні 1/3 + 2/3 - 3 самки і 6 самців.
Зміна співвідношення статей у потомстві може також викликатись і перезріванням ікри до запліднення. За значного перезрівання яєць кількість самців зростає до 100%. Виявилось, що затримка ікри у порожнині тіла самиці на різну кількість днів викликає у молоді, яка розвивалася з цієї ікри, спочатку деяке переважання самиць, а потім самців. Вважають, що затримка ікри в порожнині тіла самиці на 4-7 днів викликає її дозрівання, тому ця затримка не викликає зростання кількості самців. Перезрівання ікри більше, ніж на 21 день здійснити практично важко, оскільки самиці погано переносять це витримування, наслідком чого є низька запліднюваність ікри. Однак, перезрівання можна отримати швидше, витримуючи самицю за більш високої температури, в недостатній кількості розчиненого у воді кисню.
Мінливість і успадкованість маси і довжини тіла, часу статевого визрівання і плодючості риб, що витікає з попередньої інформації має виключне значення для розуміння процесів та явищ, які значною мірою визначають ефективність використовуємих технологій у рибництві. Для всіх видів риб характерна мінливість швидкості росту. На ріст риб впливає дуже багато генів, оскільки будь-які зміни у будові риб і в функціях тих чи інших органів може так чи інакше відбитися у першу чергу на споживанні і засвоєнні їжі. Багаточисельні чинники водного середовища, які можуть суттєво змінити інтенсивність і спрямованість метаболічних процесів, а відповідно і швидкість росту риб.
Вплив середовища на ріст риб може бути встановлений ще в період розвитку ооцитів у яєчнику самки; різні ооцити в залежності від розташування в яєчнику можуть отримувати різне забезпечення, тому ростуть з різною швидкістю і накопичують різну кількість поживних речовин. Ще більше значення має неоднаковий ріст ооцитів в момент овуляції. Після овуляції і запліднення середовище викликає додаткову диференціюючу дію на зародки риб, які знаходяться в ікринці. В період інкубації ікри також спостерігаються досить суттєві відмінності у забезпеченості ікринок киснем, у температурному і світловому режимі тощо.
Накладуючись на відмінності, які мають місце до овуляції, ця неоднорідність умов приводить до прискорення розвитку одних і уповільненню розвитку інших ембріонів, результатом чого виявляється зростаюча асинхронність ембріогенезу.
Викльов передличинок в одній і тій же порції ікри, зазвичай, розтягується на багато годин. Спадкоємність часу вилуплення для райдужної форелі невисока, а у сазана коефіцієнт варіації, як правило, не перевищує 5-6%.
Крупні і особливо ті передличинки, які виклюнулися раніше, швидше переходять до самостійного живлення і випереджають за швидкістю росту своїх ровесників. У цьому випадку варіація серед підростаючих мальків збільшується, більше може посилити цю мінливість риб харчова конкуренція.
Дещо пізніше вплив чинників середовища за достатньої кількості їжі починає згладжуватись, що приводить до поступового вторинного зниження показників фенотипової мінливості. У коропа цей процес продовжується тривалий час.
При дослідженні мінливості маси і довжини тіла у риб були встановлені певні закономірності. Розподіл риб всередині однорідної за походженням популяції або генетичної групи виявляється або близьким до нормального, або характеризується більшою чи меншою позитивною асиметрією.
Утримання коропів в умовах полікультури за надто щільних умов, коли корму для всіх видів риб зі схожим спектром живлення не вистачає, інколи стимулює різко асиметричний розподіл. Якщо з такої популяції вилучити найбільш крупних риб, то за збереження і у подальшому харчової конкуренції місце «чемпіонів» швидко займають інші екземпляри. До розтягування варіації у праву сторону приводять переваги
у живленні, які отримуються найбільш крупними особинами, які фактично віднімають корм у своїх ровесників. Збільшенню мінливості сприяє позитивна кореляція між вихідними розмірами і швидкістю росту. У риб зі зграйною поведінкою і свого роду «школами», в яких мальки наслідують один одному і не схильні до агресії в період живлення коефіцієнти варіації маси і довжини тіла зростають не так сильно, - розподіл залишається близьким до нормального. У доместикованих видів риб в результаті селекції накопичуються домінантні гени, які сприяють прискореному росту.
За вирощування одних і тих же нащадків окремо і разом коефіцієнти наслідування вище у випадку сумісного вирощування. Генетичні відмінності зростають в основному внаслідок посилення під впливом харчової конкуренції взаємодії генотипу і середовища.
Швидкість росту риб тісно пов’язана з плодючістю і життєздатністю - головними компонентами племінної цінності або відтворюваності особини. Ознаки, які визначають племінну цінність, тобто здатність залишати достатньо багато чисельне і життєстійке потомство, у всіх тварин характеризуються дуже невисокою спадкоємністю. За всіма такими ознаками безперервно відбувається інтенсивний відбір, створюються складні системи взаємопов’язаних алельних і неалельних генів, підвищується вплив неаддитивної генетичної мінливості, в результаті чого знижується спадкоємність.
Значне прискорення росту найчастіше виявляється зв’язаними з певними дефектами у розвиткові статевих залоз, які обумовлюють уповільнення або припинення статевого визрівання. Проте, у більшості випадків вони є спадковими ознаками і накопичення таких «мутацій стерильності» в результаті відбору крупних риб може мати негативний результат в селекційній роботі.
До числа ознак, які безпосередньо визначають племінну цінність риб, відносяться час вистигання плідників і їх плодючість. Спеціальні дослідження показали, що у коропа прискорене визрівання визначається багатьма, переважно домінантними генами, що не виключає наявності аномалій у інших видів риб.
Одночасно доведено, що час вистигання і нересту райдужної форелі у більшій мірі залежить від генотипу, що пропонує матеріал для продовження відповідних досліджень в цьому напрямку.
Встановлення закономірностей розвитку статевих залоз у риб, як представників нижчих хребетних тварин, має велике теоретичне і практичне значення, яке пов’язане з проблемами темпів індивідуального розвитку, системогенезу, етапності розвитку, динаміки популяцій тощо.
Швидкість розвитку репродуктивної системи у риб тісно зв’язана з умовами існування особин, а швидкість настання статевої стиглості впливає на вікову і статеву структуру нерестових популяцій і тим самим відображується на інтенсивність їх відтворення, на стабільність і цінність промислових уловів. Особливо чітко виявляються зміни у розмноженні риб і співвідношенні цінних і малоцінних видів в промислових умовах в результаті реконструкції внутрішніх водойм і господарської діяльності людини.
Вважають, що статева стиглість риб настає з моменту першого розмноження особин і закінчується припиненням функціонування статевих залоз в період старості. Зазвичай, статева стиглість у більшості видів риб настає у період переважання вагового приросту над лінійним, при досягненні особинами певного рівня обміну речовин, здатного забезпечити процес розвитку і виметування статевих клітин, багатих запасними поживними речовинами.
Необхідно підкреслити, що теорія етапності розуміється як теорія індивідуального розвитку організмів, етапи і періоди якого незворотні і послідовно змінюють один другого протягом всього онтогенезу - від початку розвитку і до смерті особини. Кожен з етапів характеризується певними морфо- еколого- фізіологічними ознаками розвитку всього організму. У кожного виду послідовність і тривалість окремих етапів і
періодів знаходяться у певній кореляції між собою, створюючи тим самим цілісний життєвий цикл.
Дослідженнями Б. В. Кошелєва встановлено, що статева залоза починає функціонувати в різні періоди й етапи індивідуального розвитку. У зв’язку з переміщенням часу першого нересту (статевої зрілості) спостерігаються зміни не лише в тривалості окремих періодів індивідуального розвитку. А саме ювенільного, дорослого організму, старості, але й у співвідношенні швидкостей розвитку всього організму й окремих органів, зокрема, статевих залоз. Тому у цих випадках не виявляються прямі зв’язки між масонакопиченням і лінійним ростом, віком особин і швидкістю розвитку відтворювальної системи за умов існування. Між тим помічено, що у багатьох випадках для розвитку організму в цілому і для росту і розвитку статевих залоз необхідні різні умови. У деяких випадках сприятливі умови для розвитку організму і, зокрема, для нарощування лінійних розмірів і маси тіла є несприятливими для розвитку статевих залоз і навпаки.
Детальне вивчення закономірностей росту у риб дозволило В. В. Васнєцову довести пристосувальне значення росту і особливості росту риб протягом всього життя особин за різних умов існування. Він відмічає, що за різних змін умов середовища межі етапів можуть змінюватись, а скоріше всього, очевидно, відбувається переміщення моменту настання статевої зрілості з одного етапу на другий.
Між тим Б. В. Кошелєв вніс деякі доповнення до теорії етапності В. В. Васнєцова. Він вважає, що послідовність адаптаціоґенезу зв’язана з еволюцією фізіологічних пристосувань організму, а в першу чергу етологія, рівень і характер обміну речовин, а потім з морфологічними змінами, а саме екстер’єр, швидкість розвитку всього організму, різна скоростиглість тощо. Лише з урахуванням специфіки обміну речовин, характеру і рівня метаболізму відображається тісний взаємозв’язок індивідуального розвитку організму з умовами існування особин і процес еволюції онтогенезу.
Продовженням аналізу пристосувального значення росту риб і виявлення зв’язку темпу росту зі швидкістю настання статевої зрілості виявились дослідження з встановлення закономірностей динаміки чисельності риб у зв’язку з забезпеченістю їжею. Узагальнення інформації з вияснення внутрішньовидових закономірностей визрівання і динаміки плодючості у риб. Ю. Є. Лапін і Ю. Г. Юровицький (1959) основну увагу приділили динаміці чисельності риб і поясненню саморегуляційного процесу, які базуються на двох основних положеннях:
- Чисельність стада риб знаходиться у відносній відповідності з кормовими ресурсами водойми;
- Статеве вистигання у риб зв’язано не з віком, а у більшій мірі з розмірами і масою тіла, тобто з темпом лінійного росту і масонакопичення.
Пряма залежність вистигання від віку характерна лише для популяцій, а не для виду в цілому. Автори показали чотири форми зв’язків між темпом росту і швидкістю статевого вистигання: 1 швидкий ріст і прискорене вистигання; 2 повільний ріст і уповільнене вистигання; 3 швидкий ріст і уповільнене вистигання; 4 повільний ріст і прискорене вистигання. При цьому відмічається, що зменшення кормності водойми до деяких меж викликає зменшення іхтіомаси популяції в її попередньому якісному стані. Зниження кормності водойм нижче деякої межі викликає вже якісну зміну особин: вони стають більш дрібними, вистигання у них відбувається вже у іншому співвідношенні з ростом. Свідченням цього явища є багаточисельні спостереження щодо біології карликових форм і різних видів риб, що характерно для плітки, карася, ляща, форелі тощо. Вивчення різних форм, які населяють одну й ту ж водойму, показує, що за низького темпу росту у карликових форм спостерігається більш раннє настання статевої стиглості. При цьому самки повільноростучих форм характеризуються високою відносною плодючістю і високою відтворювальною здатністю, порівняно із особинами із середнім темпом росту в даних популяціях. Багаточисельні дані про настання статевої стиглості. у одного й того ж виду риб у різних водоймах переконливо показують, що настання статевої зрілості може відбуватися за самої різної довжини і у різному віці.
У зв’язку з різним віком настання статевої зрілості змінюється структура нерестової частини популяції, змінюється також тривалість життя особин, що суттєво впливає на швидкість відтворення окремих популяцій.
Ці закономірності у зміні тривалості розвитку статевих залоз і швидкості настання статевої стиглості є однією з форм пристосування організму до різних умов оточуючого середовища.
Заслуговує на увагу аналіз показників росту у зв’язку з початком функціонування відтворювальної системи у окремих видів риб з раннім, середнім і пізнім настанням статевої стиглості. У всіх випадках відмічені відносно низькі показники приросту маси порівняно з довжиною риби. По мірі розвитку особин відбувається зниження темпу росту риби в довжину, що спостерігається при аналізі змін абсолютних і ще більш чітких показниках приросту довжини риби порівняно з динамікою зміни її маси. Абсолютна і відносна маса особин з віком поступово зростає і в момент, коли приріст маси, виражений у відсотках, переважає над лінійним відносним приростом риби, як правило, починає функціонувати відтворювальна система у риб.
У різних водоймах у риб, об’єктом досліджень слугував лящ, спостерігається різна інтенсивність валового приросту і швидкість падіння приросту довжини, внаслідок чого відбувається зміна часу, тобто віку, коли приріст маси починає переважати над лінійним. В процесі досліджень встановлено, що у південних водоймах тривалість цього періоду достатньо мала. За значного лінійного приросту риби протягом першого року у подальшому відбувається швидке падіння приросту довжини тіла порівняно з інтенсивним збільшенням її маси. Швидке падіння лінійного приросту особин з досить раннім і швидким зростанням її маси зв’язано з скороспілістю, з раннім досягненням особинами даної популяції термінів першого нересту. Повільне падіння лінійного приросту риби з поступовим і незначним збільшенням відносного приросту її маси зв’язано з пізніми термінами настання статевої стиглості, яка відбувається у віці 6-7 років (оз. Ільмень). На північній межі ареалу ляща (Сямозеро), не дивлячись на досить високі показники абсолютного приросту маси риби, відносний приріст як довжини, так і маси порівняно невеликий, а переважання нарощування маси над лінійним ростом у ляща спостерігається досить пізно і складає 8-9 років. У той же час відмічено перший нерест, тобто відбувається настання статевої зрілості у особин ляща у досліджених популяціях.
Аналіз деяких біологічних показників росту у риб з ранніми термінами статевої стиглості, але з одночасним вистиганням самців і самиць показує, що швидкий темп приросту довжини протягом перших років життя різко знижується після настання статевої стиглості, а приріст маси особин досить швидко починає переважати над лінійним, що погоджується з періодом настання термінів першого розмноження.
При порівнянні показників росту самиць і самців бичка-кругляка в Азовському морі і відсоткового співвідношення вікових груп в промислових уловах виявилось, що не дивлячись на більш високий темп росту самців, статева стиглість у них настає на один рік пізніше, ніж у самиць. За різного віку настання статевої стиглості у самиць у віці двох років і самців на один рік пізніше, у них виявлена різниця як у розмірах, так і в темпі валового приросту і швидкості падіння лінійного росту особин. У зв’язку з відміченими особливостями в рості і віці статевозрілих самців і самиць спостерігається різночасовий підхід до місць розмноження одновікових особин однієї статі, де відбувається промисловий інтенсивний лов бичка-кругляка. Цим визначається неоднакове значення окремих вікових груп в особливостях процесу, а саме - відціджуючі знаряддя лову вилучають зі стада 74% швидкоростучих самців і біля 64% самиць дворічного віку.
Аналіз літературних джерел щодо специфіки росту риб переконливо свідчить про те, що у багатьох випадках швидкість статевого вистигання не має прямого зв’язку ні з віком, ні з довжиною, ні з масою особини. Однак, очевидно, для окремих популяцій за відносно стабільної кормової бази та екологічних умов існування може спостерігатись пряма залежність між темпом росту і часом першого ікрометання. Лише за відносно стабільних умов середовища для особин даної популяції умови розвитку можуть виявитися сприятливими для вагового і лінійного росту, а також для розвитку відтворювальної системи.
Узагальнення багаторічних матеріалів досліджень щодо існуючих в природі співвідношень між темпом росту особин і швидкістю досягнення термінів першого ікрометання показано, що лише в деяких випадках спостерігається пряма залежність між темпом росту риби і швидкістю настання статевої стиглості. Якщо зростає темп росту, то відбувається досить раннє визрівання особин, а якщо темп росту падає, то відбувається більш пізнє настання статевої стиглості риб. Однак у багатьох випадках, не дивлячись на високий темп розвитку особин, спостерігається пізнє вистигання самців і самиць, а за низького темпу росту самок і самців, навпаки, спостерігається у них більш раннє настання статевої стиглості. Різний темп росту з неоднаковою швидкістю настання статевої стиглості можна виявити у самців і самиць різних видів риб. У більшості випадків темп росту самиць і самців однаковий і настання статевої зрілості спостерігається у них в одному віці. У деяких випадках темп росту самиць і самців може бути різним. Інколи може спостерігатися високий темп росту самиць за раннього настання статевої стиглості порівняно з самцями, у яких може бути низький темп росту і пізнє вистигання. У ряді випадків за високого темпу розвитку самців їх статева зрілість настає у більш пізньому віці, ніж у самиць, у яких, не дивлячись на низький темп росту, реєструється раннє настання статевої стиглості.
Отже, існування прямого зв’язку між темпом росту і настанням статевої стиглості у риб спостерігається лише у тих випадках, коли умови
існування популяцій відносно стабільні, не виходять за межі звичайних для даного виду і є сприятливими як для розвитку всього організму і, зокрема, для лінійного і вагового росту особин, так і для темпу розвитку відтворювальної системи. За інших екологічних умов, які досить часто значно відхиляються від типових, що спостерігається під впливом господарської діяльності людини і зміни зв’язків, які історично склались між організмом і водним середовищем у ряду прісноводних видів риб, прямий зв’язок між темпом росту і швидкістю статевої зрілості може порушуватись. Тоді на перший план в якості однієї з форм адаптації популяції виступає зворотній зв’язок між цим процесами, коли особини визрівають швидше за низького темпу росту. Подібного роду зв’язки відмічені у видів з різною екологією і, зокрема, з розмноженням, і є своєрідними формами адаптаціогенезу. За глибоких і незворотних змін умов існування, а також на межах поширення виду, у різних екологічних форм і за статевого диморфізму спостерігається неодночасність у розвиткові окремих органів, і, зокрема, відтворювальної системи, що, безперечно, зв’язано із зміною обміну речовин у самок і самців і є пристосуванням особин, екологічних форм і популяцій до різних умов існування.
Виходячи з розглянутої концепції, складові статевої стиглості і вистигання статевих продуктів мають безпосереднє теоретичне і практичне значення. Розробка заходів, спрямованих на підвищення рибопродуктивності водойм шляхом управління рибничими процесами вимагає глибоких знань життєвого циклу промислово цінних видів риб. Методи управління в основному базуються на оптимізації процесів розмноження риб.
Відомо, що розмноження - це ланка життєвого циклу риб, яка забезпечує у взаємодії з іншими ланками відтворення популяції і збереження виду. Для кожного виду риб характерні свої специфічні особливості розмноження, які обумовлені адаптацією риб до певних екологічних умов розмноження і розвитку молоді, яка дає поповнення, необхідне для збереження виду і підтримання його чисельності. Поняття розмноження включає і розвиток статевих залоз, нерест, осіменіння, запліднення, ембріональний і постембріональний розвиток.
Промислово цінні види риб, які населяють певні водойми, є, зазвичай, різностатеві. Статеві залози самок називаються яєчниками, у них утворюються зрілі статеві клітини - яйця (ікринки). Статеві залози самців називаються сім’яниками, у них утворюються зрілі чоловічі статеві клітини - сперматозоїди.
Статева стиглість у окремих видів риб настає у різному віці. Час настання статевої стиглості у одного й того ж виду може бути також різним. Він варіює не лише у різних популяцій одного й того ж виду, але й у риб однієї популяції, природного ареалу.
Це явище розглядають як адаптацію організму риб до умов середовища з метою найбільшого забезпечення відтворення виду.
Більшість коропових, окуневих і лососевих риб досягають статевої зрілості у віці 2-4-х років. У деяких видів риб період розвитку статевих клітин затягується на більш тривалий період. Осетрові стають статевозрілими у віці 6-12 років, при цьому статева стиглість у самців, зазвичай, настає на 1-2 роки раніше, ніж у самиць.
Розвиток статевих клітин може прискорюватись або уповільнюватися під впливом чинників водного середовища, а у деяких випадках навіть призупинятися. Лящ, який населяє наші південні водойми, стає в масі статевозрілим на 3-4 році життя. У водоймах центральних водоймі він визріває на 4-5-му році життя. У північних водоймах з порівняно низькою річною температурою статева стиглість у ляща настає на 5-7-му році. Аналогічна швидкість вистигання статевих клітин спостерігається і в інших видів риб і це не явище а закономірність.
Встановлено, що низькі температури призупиняють кінцеві стадії визрівання статевих продуктів у осетрових риб. Недостатнє живлення риб стримує вистигання статевих продуктів. Між тим фізіологічно повноцінне живлення, яке повністю забезпечує організм в поживних речовинах, прискорює їх вистигання. Зменшення чисельності популяції риби під впливом промислу приводить до підвищення забезпеченості їжею, а це відображується на прискоренні статевого вистигання особин.
Зсуви у часі настання статевої стиглості у риб можуть відбуватися під дією чинників зовнішнього середовища не лише у різних популяцій одного й того ж виду, але й у однієї і тієї ж популяції; волжська білуга, досягає статевої стиглості у віці від 10 до 18 років.
Розвиток жіночих статевих клітин (оогенез) і чоловічих статевих клітин (сперматогенез) є досить тривалим і складним процесом. Кожна статева клітина, перш ніж вона повністю визріє, повинна обов’язково пройти у своєму розвиткові відповідні стадії. При цьому розрізняють дві складові: - період до досягнення статевої стиглості, починаючи від виникнення первинних статевих клітин і, закінчуючи утворенням зрілих статевих продуктів; - періодичне вистигання певної частини статевих продуктів протягом міжнерестового періоду після досягнення статевої стиглості.
Перший період є більш тривалим, порівняно з другим, і у різних видів риб він займає неоднаковий час. Періодичне ж визрівання статевих продуктів у статевозрілих особин вимагає меншого часу, але знову ж-таки тривалість його у різних видів неоднакова. Сазан, лящ, судак і багато інших видів риб розмножуються, зазвичай, щорічно, а осетрові риби лише через 3-5 років, а окремі особини лише через 9 років, виключенням є тихоокеанські лососі, які після розмноження гинуть.
Стадії стиглості статевих залоз можна визначити за допомогою розроблених для кожної групи риб так званих шкал стиглості, які відображають специфічні особливості процесу визрівання статевих продуктів лише у даної групи риб.
О. Ф. Скакун, Н. А. Буцкол розробили дві універсальні шкали для наших промислових груп риб. Перша шкала відображує загальний хід визрівання статевих продуктів у самиць, а друга - у самців. Ці шкали дозволяють визначити стадії стиглості статевих залоз за їх зовнішнім
виглядом і гістологічною будовою. На основі цих двох шкал розроблена єдина універсальна шкала зрілості статевих залоз самок і самців.
Попередня інформація, яка викладена у відповідному розділі, переконливо свідчить про умовну універсальність розглядаємого процесу, але у різних видів риб, на фоні загальної закономірності простежуються відповідні видоспецифічні особливості, що необхідно враховувати при формуванні ремонтно - маточних стад.
Попередня інформація переконливо свідчить про наявність відповідного зв’язку між швидкістю настання статевої стиглості з умовами існування, віковою структурою і динамікою чисельності популяцій.
Відомо, що зарегулювання стоку рік, створення водойм - охолоджувачів ТЕС, здійснення заходів з акліматизації гідробіонтів тощо приводить до значних змін у фауні внутрішніх водойм, які проявляються у зміні видового складу фауни риб, їх морфо-фізіологічних характеристик, швидкості росту та настання статевої стиглості. Наслідком всіх цих заходів та змін є поява у водоймах карликових форм і риб-«гігантів» з різною швидкістю розвитку репродуктивної системи.
Існує безліч факторів переміщення термінів настання статевої стиглості на різні етапи і періоди індивідуального розвитку організму. Детально описані випадки того, що самці лосося нерідко стають статевозрілими в річці, не виходячи в море. Вони можуть мати зрілі статеві продукти за довжини всього 10 см, не переходячи в період дорослого організму. Між тим, самиці такої ж довжини, які населяють ту ж річку, мають зовсім недорозвинені яєчники.
Самці прохідних видів риб, що рано розвиваються і не покидають нерестових рік, суттєво відрізняються від дорослих прохідних самців не лише розмірами, а й цілою низкою морфологічних ознак. У далекосхідних лососів не виростають і не викривляються челюсті, не виростають крупні зуби, окрас тіла зберігає схожість з пестрятками, становлячись лише дещо яскравішим. У них майже не змінюються покриви тіла і не спостерігається типової картини дегенерації кишківника, як це має місце у прохідних
самців. Не дивлячись на затримку у розвитку всього організму карликових самців у них десь на стадії пестрятки починає функціонувати відтворювальна система, тобто відбувається зміщення статевої стиглості на більш ранній період індивідуального розвитку.
Аналогічні зміни у розвиткові відтворювальної системи спостерігається і в інших видів риб. Не дивлячись на уповільнений темп росту і затримку в розвитку на більш ранніх етапах онтогенезу, у тугорослого карася спостерігається відносно швидкий розвиток відтворювальної системи. Деякі зміщення початку функціонування відтворювальної системи на більш ранні етапи індивідуального розвитку є одним із способів пристосування особин даного виду до різних умов існування. У ляща, який населяє південні райони, статева зрілість настає у ранньому віці за своєрідного екстер’єру плідників порівняно з особинами в популяціях, що населяють водойми поблизу північної зони ареалу цього виду. За великої довжини тіла крупні і високотілі лящі північних водойм досягають статевої зрілості у 9-10-ти річному віці, а статева зрілість у ляща, який населяє південні водойми настає у дволітньому віці за довжини тіла всього 5,5 см. Порівняльний аналіз форми тіла на ранніх етапах розвитку ляща з формою тіла карликового ляща переконливо свідчить про те, що у карликової форми відбулася затримка в розвиткові екстер’єру на ранніх етапах онтогенезу. Не дивлячись на розвиток відтворювальної системи у карликової форми розвиток відбувається у швидкому темпі і особини досить рано починають розмножуватися, це явище спостерігається у віці 2-3 роки, але тривалість життя особин у таких популяцій скорочується до 5-6 років.
Отже, початок функціонування відтворювальної системи у риб може спостерігатися на різних етапах і періодах індивідуального розвитку. Природно, що у таких випадках буде мати місце нерівномірна швидкість розвитку організму і репродуктивної системи, а час настання статевої стиглості у особин різних популяцій не буде зв’язаний ні з віком, ні з довжиною, ні з формою тіла. Ці закономірності різночасовості розвитку відтворювальної системи і форми тіла протягом окремих етапів і періодів індивідуального розвитку є одним із шляхів адаптації організму до різних умов існування, тобто шляхом розширення ареалу даного виду.
Відомий вчений І. І. Шмальгаузен (1935) відмічав, що процес росту не можна собі уявити ізольованим від розвитку, а організм, що розвивається, не зв’язаним з тим середовищем, в якому відбувається процес. Характер розвитку і росту є специфічним для кожного виду, а потенційні можливості зміни розвитку і росту за зміни умов існування мають різну варіабельність і проявляються у різній формі. Вивчаючи особливості росту риб, які населяють різні географічні широти, Т.С. Расс (1948) відмічав, що специфіка й розвиток навіть у близькородинних видів безпосередньо зв’язана з різною тривалістю проходження окремих фаз і стадій протягом індивідуального розвитку. Вивчаючи процеси онто- і філогенезу О. М. Северцов (1949) вказував, зокрема, на відносно пізнє закладання репродуктивної системи у ссавців, пояснюючи це тим, що функціонування органів відтворювальної системи, яка передбачає відтворення нащадків і піклування про них батьків, повинно настати у той період індивідуального життя особин, коли за сумою біологічних умов батьки досягають повної сили і здатні прогодувати і захистити підростаючих нащадків.
У ікромечущих видів риб, всі стадії розвитку яких відбуваються у водному середовищі, онтогенез, а також гонадо- і гаметогенез у більшій мірі, ніж у теплокровних, зв’язані з тими або іншими абіотичними чинниками, оскільки у них інтенсивність процесів обміну речовин і діяльність залоз внутрішньої секреції залежить, від тієї чи іншої температури води та інших чинників. Тому і нейрогуморальна регуляторна система є наче б «відкритою системою», діяльність якої знаходиться у більшій залежності від безпосереднього впливу умов існування, ніж у гомойотермних тварин. Основні біохімічні та фізіологічні процеси, які зв’язані з розвитком, ростом, обміном речовин і розмноженням риб, узагальнюються і контролюються у більшій чи меншій мірі гормонами.
Однак, ендокринний контроль риб, які розмножуються щорічно, повинен у значній мірі відрізнятися від ендокринного контролю у ссавців, які мають досить складний статевий цикл. Існує інформація про те, що до цього часу жоден дослідник не зміг добитися тривалого росту ооцитів у пойкілотермних тварин від первинних овоцитів до повністю розвиненої яйцеклітини за допомогою фолікул-стимулюючого гормона. Порівняно з іншими хребетними тваринами у риб розвивається значна кількість статевих клітин. Коефіцієнт зрілості, який характеризується відношенням маси статевих клітин до маси тіла, що виражений у відсотках, становить у різних видів риб перед ікрометанням у середньому біля 20%. У овоцитах, що розвиваються, накопичується велика кількість поживних речовин. У видів з осінньо-зимовими термінами накопичення поживних речовин в овоцитах, процес гаметогенезу і, зокрема, період інтенсивного вітелогенезу, має тривалий проміжок часу, який визначається шістьма місяцями. Вказані особливості розвитку статевих клітин у риб, зв’язані з накопиченням значної кількості поживних речовин в овоцитах, що розвиваються, з великою плодючістю, з тривалим періодом вітелогенезу, адекватно умовам існування, а саме впливу пресу хижаків, холоднокровністю, морфо-екологічними особливостями всіх періодів онтогенезу тощо. Тривалість перебігу окремих етапів і періодів онтогенезу, різночасовість розвитку органів, систем органів і всього організму в цілому тісно зв’язані екологією окремих видів в цілому і з багатьма екологічними чинниками, які впливають на характер перебігу цих процесів.
Встановлено, що багато факторів середовища абіотичних та біотичних складових можуть впливати на специфіку й інтенсивність обміну речовин як у самиць, так і у самців, зміна якого, у свою чергу, приводить до зміни корелятивних співвідношень в темпі розвитку окремих органів, впливає на екстер’єрні особливості особин у зв’язку з різночасовістю розвитку організму, його массонакопичення і лінійного росту зі швидкістю розвитку окремих органів і в першу чергу статевих залоз.
Отже, початок функціонування відтворювальної системи у риб переміщується на різні етапи й періоди онтогенезу і є одним із способів адаптації особин до різних умов існування. Початок функціонування статевої залози, тобто часу першого нересту риб, спостерігається, зазвичай, на початку періоду дорослого організму, або статева стиглість може зміщуватися на більш ранні або більш пізні етапи і періоди індивідуального розвитку. Інколи статева залоза починає функціонувати у особин вже в мальковий період. У деяких випадках відбувається затримка у темпі розвитку гонад, а початок функціонування відтворювальної системи, тобто перше розмноження особин спостерігається в середині періоду дорослого організму, і на кінець, статева зрілість може наставати досить пізно. Слід зазначити, що у особин з пізнім настанням статевої стиглості значно подовжується тривалість життя плідників у популяціях, порівняно з самицями і самцями з ранньою статевою стиглістю, з раннім досягненням термінів першого нересту.
Таким чином, у більшості випадків у різних популяцій видів риб з широким ареалом відсутні прямі зв’язки між темпом росту і віком особин, із швидкістю розвитку і початком функціонування відтворювальної системи. Зазвичай, у більшості риб спостерігається інтенсивний лінійний приріст в ювенільний період, а інтенсивний приріст маси - в період дорослого організму. В період переважання відносного приросту маси особин по відношенню до їх довжини, коли відбувається начеб-то «перехрест» цих величин, настає статева стиглість у риб, тобто спостерігається перший нерест. Між тим біологічні показники росту риб піддаються великим змінам як у окремих видів, які населяють різні зони ареалу, а саме в центрі і на його південній і північній межах, так і у різних екологічних форм. Все це приводить у ряді випадків до неодночасності і розходження в швидкості росту маси тіла і лінійного росту, темпі розвитку репродуктивної системи. Темп розвитку відтворювальної системи у риб тісним чином зв’язаний з рівнем і характером метаболізму в особин у зв’язку з різними умовами існування.
Зміна в характері і рівні обміну речовин викликає ту чи іншу різночасовість у розвиткові всього організму - його рості, формі тіла, і швидкості настання статевої стиглості, яка зв’язується з початком функціонування статевих залоз, тобто з першим нерестом.
Зміна швидкості розвитку відтворювальної системи і часу досягнення статевої зрілості зв’язано з різночасовістю розвитку всього організму і окремих органів. Тому початок функціонування статевих залоз може бути приурочено до того або іншого етапу і періоду індивідуального розвитку.
При цьому всі зміни є реакцією організму на зміну умов існування і одним із способів адаптації особин до різних умов середовища. Віку статевого визрівання присутня різна індивідуальна мінливість. У окремих популяцій за зміни умов існування відсоток визріваючих самиць, які відносяться до різних поколінь, має дещо змінюватися, тобто спостерігається різна швидкість настання статевої стиглості. У ранніх вікових груп, що у деяких випадках суттєво впливає на структуру всього нерестового стада, значно змінюючи величину поповнення і залишку. Всі ці зміни швидкості настання статевої стиглості у особин і окремих популяціях за відносно стабільних умов їх існування має певний розмах варіабельності і спостерігається пряма залежність статевої стиглості від умов існування, яка зв’язана, досить часто, з темпом росту особин, то у виду в цілому зміни у термінах настання статевої стиглості мають більш складний характер, який зв’язаний із специфікою розвитку всього організму і відтворювальної системи. Зміни у тривалості розвитку статевих залоз у самок зв’язані з подовженням або скороченням термінів проходження ІІ стадії стиглості яєчників, періоду протоплазматичного розвитку овоцитів, кінцеві фази якого характеризуються овоцитами у фазі одношарового фолікула. Прискорення або уповільнення швидкості розвитку статевих залоз у самиць, які досягли терміну першого нересту,
припадає в основному на період превітелогенезу. У зв’язку з різним темпом досягнення термінів статевої стиглості змінюється і структура популяцій: за раннього настання статевої стиглості нерестова частина популяції складається з молодших вікових груп, граничний вік життя у таких популяціях невеликий. У більшості випадків це другий тип нерестової популяції, коли залишок менше поповнення. За пізнього настання статевої стиглості у особин нерестова частина популяції складається з великої кількості різновікових поколінь - це третій тип нерестової популяції, коли залишок переважає над поповненням. Особини у таких популяціях приймають участь у розмноженні багато разів протягом всього життя. Граничний вік життя плідників у таких популяціях досить великий. Природно, що швидкість відтворення популяцій з раннім терміном настання статевої стиглості у особин з коротким віковим рядом нерестової частини популяції і тривалим життям особин, значно вище, ніж у популяцій з пізнім терміном настання статевої стиглості у особин, з розтягненим і великовіковим складом плідників у статевозрілій частині популяції, з відносно тривалим періодом життя особин в таких популяціях. У зв’язку з глибокими і незворотними змінами умов існування, популяції першого типу виявляються менш вразливими і досить швидко відновлюють свою чисельність.
Популяція другого типу, не дивлячись на велику стійкість до короткочасних змін умов існування, дуже важко і протягом тривалого часу відновлюють свою чисельність за тривалих несприятливих умов існування, і у тому числі необхідних для перебігу всіх ланок репродуктивного циклу.
Особливості розвитку ооцитів у риб до настання термінів першого ікрометання - теоретична складова яка значною мірою визначає технологію відтворення в штучних умовах, що є вагомою складовою рибництва.
В іхтіологічній літературі досить поширена інформація про різний час настання у риб статевої стиглості. Зміни у тривалості проходження стадії стиглості статевих залоз у самиць в період досягнення термінів
першого нересту припадає на початкові стадії розвитку гонад (І і ІІ). Відомо також і те, що тривалість ювенільного періоду досить різна не лише у різних видів риб, але вона достатньо схильна до стійких змін у одного і того ж виду, поширеного у водоймах різних широт з різним гідрологічним режимом. У зв’язку з цим серед прісноводних костистих риб можна виділити скороспілі і пізньоспілі популяції певних видів. Виходячи з цього, в інтересах рибництва тривалість цього процесу можна змінити.
Вважають, що управління онтогенезом у риб в рибогосподарських цілях, очевидно, буде полягати у створенні оптимальних умов для кожної фази розвитку, а також у зміні режиму розвитку, у прискоренні або, навпаки, уповільнені його, збільшенні або, навпаки, скороченні тривалості тієї чи іншої фази, а звідси і в зміні типу і характеру всього життєвого циклу.
Отже, вивчення специфіки перебігу стадій стиглості статевих залоз і особливостей розвитку ооцитів у риб в період настання часу першого ікрометання має не лише велике теоретичне, але й практичне значення.
Дослідженнями Б. В. Кошелєва встановлено, що у різних видів риб у зв’язку зі зміною умов існування спостерігаються значні зміни у тривалості проходження періоду протоплазматичного росту ооцитів. Ці фази стиглості овоцитів характерні для ІІ стадії стиглості статевих залоз. Саме в цей період розвитку статевих клітин відбуваються найбільш суттєві зміни в ядерно-плазмових співвідношеннях в овоциті, що розвивається. Ці зміни є попередниками наступного періоду, зв’язаного з накопиченням поживних речовин в овоцитах.
В процесі ово- і сперматогенезу є різні періоди, протягом яких можуть відбуватися тривалі затримки у розвиткові статевих залоз у самців і самиць. Затримки у розвитку статевих клітин у самиць відмічалося після анатомічної диференціації гонад, що характеризувалося завершенням формування яйценосних пластинок, в яких овоцити знаходилися на різних фазах стиглості періоду протоплазматичного росту. Саме в цей період у самиць лососевих можливі тривалі затримки у розвиткові відтворювальної системи, які припадають на період превітеллогенезу аж до початку трофоплазматичного росту ооцитів. У самців лососевих риб розглянуті затримки можуть відбуватися в той період розвитку гонад, котрий характеризується збільшенням числа статевих клітин у сім’янику і продовжується анатомічною диференціацією, а саме формування ампул до цитологічної диференціації сім’яника.
Тривалі зміни у статевих циклах у риб припадають на період протоплазматичного росту ооцитів як у особин за досягнення термінів першого ікрометання, так і у плідників в період дорослого організму.
Зміни у швидкості розвитку гонад, які пов’язані з тривалістю перебігу фази протоплазматичного росту ооцитів (превітеллогенезу), приводять до збільшення або зниження часу віку настання статевої стиглості, тобто до утворення швидкоспілих або пізньоспілих форм.
Різна швидкість настання статевої стиглості має великий вплив на вікову структуру популяції, змінюючи співвідношення поповнення і залишку, початок розмноження особин, тривалість життя, кількість статевих циклів, а це, у свою чергу, без сумніву, змінює (збільшує чи зменшує) швидкість відтворення окремих популяцій. Порівняльний аналіз специфіки перебігу окремих стадій стиглості статевих залоз у риб, які досягли термінів першого нересту і розмножуються повторно, показав, що найбільш тривалі зміни у тривалості періоду протоплазматичного росту, а це ІІ стадія стиглості статевих залоз, відмічені у період досягнення термінів першого нересту. У плідників, які розмножуються другий, третій і наступні рази, ці зміни у тривалості перебігу періоду протоплазматичного росту піддаються менш значним коливанням. Взагалі тривалість ІІ стадії стиглості гонад у самок, що розмножуються повторно, значно скорочена порівняно з ІІ стадією у особин в період досягнення термінів першого ікрометання.
Для забезпечення нормального і ритмічного функціонування відтворювальної системи також необхідні певні енергетичні резерви в організмі, які б могли бути своєчасно мобілізовані для накопичення значної кількості запасних поживних речовин в овоцитах, що розвиваються. Особинами в період досягнення термінів першого ікрометання для накопичення резервних поживних речовин необхідно значно більше часу, ніж дорослим плідникам з режимом розмноження, який вже сформувався та встановився раніше. Тому у особин одного й того ж виду, які населяють водойми різних широт в межах ареалу, спостерігаються більш суттєві зміни у тривалості ІІ стадії стиглості статевих залоз, або іншими словами тривалості періоду превітеллогенезу у ювенальних особин, у яких розглядаєма стадія розтягується на декілька років, ніж у плідників за повторних статевих циклів. Тривалість ІІ стадії у певних видів риб за щорічного нересту зростає на один або декілька місяців.
Якісні та кількісні критерії обміну речовин в організмі риб безсумнівно впливають на їх ріст та розвиток. Виходячи з цього, суттєвий інтерес викликає зв’язок обміну речовин зі швидкістю розвитку статевих залоз у риб.
Розвиток статевої залози - це процес, перебіг якого відбувається у єдності зовнішнього і внутрішнього, який залежить як від попереднього стану, так і від тих умов, у яких знаходиться організм у процесі вистигання статевих залоз, розвиток яких нерозривно зв’язаний з загальним обміном організму.
У зв’язку з цим при розроблені теоретичних аспектів рибництва велика увага приділяється дослідженню обміну речовин, встановленню взаємозв’язків між специфікою обміну речовин у риб і початком функціонування репродуктивної системи. Виявлена залежність між плодючістю і швидкістю розвитку яєчників, відмічена фізіологічна неоднорідність окремих поколінь, яка впливає на темп вистигання особин в популяціях, встановлено тісний взаємозв’язок між віком плідників і якістю статевих продуктів, а також зареєстрована специфіка обміну речовин у окремих видів і своєрідність характеру метаболізму у риб з різною біологією у зв’язку з розвитком статевих залоз. Всі ці та інші дослідження розширюють нашу уяву щодо фізіологічних механізмів зміни швидкості і послідовності перебігу окремих етапів у розвитку риб і темпі розвитку окремих органів протягом всього онтогенезу.
Слід зазначити, що у риб період накопичення запасних поживних речовин в овоцитах є досить тривалим і напруженим періодом у розвитку статевої залози і особин в цілому, оскільки з ікрою в період нересту «втрачається» біля 25% маси тіла, що об’єктивно - маса статевих залоз відносно маси тіла становить біля 15-25%. Якщо в статевих клітинах, що розвиваються, як правило, протягом тривалого часу, який складає біля 4-6 місяців, накопичується велика кількість запасних поживних речовин, то природно, що для здійснення процесу вітелогенезу необхідні великі енергетичні витрати з боку материнського організму. У зв’язку з цим в організмі повинні бути відповідні резерви, які б могли бути спрямовані на розвиток великої кількості ооцитів у статевій залозі, особливо у особин в період досягнення термінів першого ікрометання. У деяких видів риб, в міру надходження поживних речовин, відбувається паралельний розвиток всього організму, що супроводжується збільшенням маси та лінійних розмірів статевих залоз. Одночасно в яйцеклітинах відбувається інтенсивне накопичення поживних речовин. У інших видів риб спочатку відбувається інтенсивний лінійний і особливо ріст маси самиць, і лише після накопичення певних енергетичних резервів починається досить швидкий розвиток ооцитів, який зв’язаний з проходженням періоду трофоплазматичного росту. Якщо за якихось причин знижується рівень і швидкість обміну речовин, то не відбувається накопичення резервних речовин в овоцитах, які розвиваються, і спостерігається затримка в їх розвиткові на фазах превітелогенезу. Однак великих енергетичних витрат вимагає не лише розвиток овоцитів, але й сам процес розмноження особливо у видів, які починають переднерестову міграцію далеко від місць ікрометання. У цьому плані досить цікавими є спостереження за нерестом морського менька, самиці якого після ікрометання деякий час знаходяться у стані спокою, потім починається період інтенсивної годівлі, а через деякий час вони знову виметують чергову порцію стиглих ікринок. На відміну від видів риб з асинхронним ростом ооцитів і порційним типом ікрометання, у морського менька тривалий нерестовий період забезпечується не одночасною, а переривистою овуляцією якоїсь частки ооцитів, яка синхронно розвивається, і які знаходяться на завершальних і однакових фазах періоду вітелогенезу.
Отже, всі зміни умов існування тісним чином зв’язані зі зміною фізіологічного стану особин і у першу чергу, зі зміною обміну речовин, що є пристосуванням організму до різних умов існування. Ці закономірності дають змогу зрозуміти зміни, які відбуваються у швидкості розвитку окремих органів в онтогенезі, розвиток яких може інколи відбуватися з різною швидкістю, змінюючи процес системогенезу.
Багатовіковий досвід людства відносно взаємозв’язку між якістю плідників і нащадків загальновідомий. Одночасно з цим характер впливу і механізм процесу вимагає певного тлумачення і теоретичного обґрунтування.
У процесі формування промислового стада риб та розроблення методів штучного відтворення цінних видів риб досить важливим є питання щодо взаємозв’язку між якістю батьків і якістю отриманих від них нащадків, а також питання щодо впливу їх життєстійкості на формування наступного покоління риби. Існує інформація щодо впливу віку плідників на якість нащадків, згідно якої від середньовікових плідників отримують кращих за якістю нащадків. Поряд з цим відомо, що самки форелі, які характеризуються високим рівнем обміну речовин, продукують нащадків також з найкращими показниками метаболічних процесів і інтенсивним ростом, особливо на ранніх етапах життя.
Між тим експериментально доведено, що личинки тарані і ляща з більшою швидкістю ембріонального розвитку характеризувались більш низькою життєстійкістю. Це пов’язують з тим, що швидкість розвитку до певної міри характеризує швидкість обміну речовин у ембріонів і загальні процеси життєдіяльності, які визначають якість даного потомства. Тому зрозумілим стає те, що у личинок, які не отримують їжі ззовні, тривалість життя буде в загальному меншою у тих, які характеризувались підвищеною швидкістю розвитку. Для самиці чотирирічної тарані, які характеризувалась найбільш інтенсивним ростом, голодуючі личинки виявились найменш життєстійкими, а у самиці ляща семирічного віку з модальним приростом виявились найменш життєстійкими личинки. Припускають, що ці зв’язки можна розуміти так, що чотирирічні самиці тарані із швидким ростом і семирічні самиці ляща з середнім ростом характеризувались найкращим обміном речовин, який і передали своїм нащадкам. Однак останні завдяки інтенсивному обміну речовин і розвивались, і закінчували своє існування за умов голодування значно швидше, ніж личинки, які отримали у спадок гірший обмін.
Особливої уваги заслуговують дані щодо впливу вмісту жиру у м’язах і на внутрішніх органах риб на життєстійкість личинок. Виявилось, що у самок ляща із збільшенням кількості жиру у м’язах і на внутрішніх органах життєстійкість личинок, які живились, в середньому зростала. У тарані, навпаки життєстійкість личинок від самиць з відносно більшим вмістом жиру у м’язах і на внутрішніх органах була меншою. Однак звертає увагу той факт, що вміст жиру у самиць ляща перевищував у 3-4 рази, ніж у тарані. Це свідчить, що жировий обмін у цих видів риб відбувається по-різному. Вважають, про те, що самці ляща на період розмноження необхідно мати більше запасного жиру, ніж самці тарані. Тому, очевидно, вміст жиру у ляща у більшій мірі відображує конституціональні властивості самиць, а звідси і життєстійкість личинок у них позитивно зв’язана з кількістю жиру у самиць. На відміну від ляща, у самиць тарані великий залишок жиру після вітелогенезу і зимівлі може означати, очевидно, порушення жирового обміну, що може свідчити про гірші конституційні властивості самиці, тому і життєстійкість личинок від таких самиць виявилась низькою.
Вивчення впливу віку на якість нащадків показало, що вік самиць впливає на ступінь заплідненості ікри і виживаність ембріонів. Найкращий результат дають самиці у розквіті життєвих сил, у стиглому віці (чотирьох - шестирічного віку), гірший - молоді самиці трьохрічного віку і найгірший - старі самиці семи-восьми років. Вік самців також впливає на запліднюваність ікри і виживаність ембріонів, але цей вплив не дуже великий. Між тим виявлено, що запліднення ікри і виживаність ембріонів у зріловікових самців чотирьох - п’яти років краще, ніж у молодих двох - трьох років і старих у віці шести-семи років.
За різного віку підбору плідників з метою штучного осіменіння з подальшим заплідненням ікри найкращий результат з запліднення ікри і виживаності ембріонів дає поєднання статевостиглих самиць і самців між собою, найгірший - поєднання старих плідників з молодими і між собою. Застосування зріловікових плідників у поєднанні з молодими і старшими плідниками покращує результат запліднення ікри і виживаності ембріонів порівняно з найгіршими поєднаннями.
Між тим доведено, що за різних вікових поєднань плідників по мірі розвитку нащадків відносна сила впливу віку плідників на виживаність нащадків в дослідних умовах поступово слабшала за відносного зростання впливу чинників середовища.
В ланцюгу робіт з розведення риб вагомою складовою є об’єктивне прогнозування впливу швидкості росту плідників на виживаність і чисельність нащадків у риб.
У більшості видів риб показники якості ікринок із збільшенням швидкості росту і плодючості самок знижуються. Між тим розмірам, масі і жирності ікринок надається виключно важливе вирішальне значення для виживання поколінь риб, оскільки вони визначають величину запасу поживної речовини в найбільш вирішальний період життя поколінь - період змішаного живлення личинок. У зв’язку з цим важливим є зв’язок між цими показниками якості ікринок і виживаністю нащадків.
Експериментальним шляхом встановлено, що тривалий період виживання голодуючих личинок, виведених із крупних і більш важких ікринок, не завжди свідчить про те, що такі ікринки є найбільш високоякісними. Тривалість життя голодуючих личинок незалежно від розмірів ікринок, з яких вони отримані, зв’язана негативним зв’язком з інтенсивністю обміну речовин. Скоріш гинуть ті з них, які мають кращий обмін, тому без їжі вони швидше витрачають запаси енергетичних речовин в тілі. Личинки, які швидше гинули за голодування, можуть виявитися за умов нормального живлення більш високоякісними, ніж ті, що тривалий час живуть без їжі.
Значно більш тісний взаємозв’язок встановлено між виживаністю личинок тарані і ляща і вмістом загального азоту, вуглеводів, кальцію і жиру в ікринках. Тривалість життя голодуючих личинок обох видів риб по мірі збільшення вмісту загального азоту в ікринках значно знижувалась, а із збільшенням вмісту вуглеводів і кальцію - в загальному зростала.
Аналогічний зв'язок із вмістом жиру в ікринках за характером був майже такий. У личинок, які живились, виживаність була найвищою за середнього вмісту в ікринках загального азоту, вуглеводів і жиру. Такий же зв’язок виявляється у виживаності личинок тарані з вмістом кальцію, а у ляща з зростанням кальцію виживаність дещо збільшувалась.
Про значення розмірів ікринок для виживання риб на ранніх етапах розвитку в літературі існують різні точки зору. Проте не виключено, що у деяких дослідах, які показали кращу виживаність нащадків із крупних ікринок, останні були отримані від зріловікових самок, які, як відомо, дають кращих нащадків. Інші автори, які вивчали вплив віку плідників на виживаність нащадків у риб, вважають, що величина ікринок не може слугувати показником їх життєздатності. Остання на думку цих вчених залежить від більш глибоких властивостей і не лише (і, може бути, не стільки) від величини запасу енергетичних і пластичних речовин в ікринці, але й від біохімічних і фізіологічних процесів, які забезпечують обмін і морфогенез ембріону, що розвивається. А величина, маса і жирність ікринок є лише слабким відображенням цих властивостей. Виключенням можуть бути лише випадки, які досить характерні, коли вони забезпечують гідростатичні властивості пелагічних ікринок певних видів риб.
Надавати вирішальне значення масі і жирності ікринок у виживанні покоління, яке визначається в період змішаного живлення, значить не визнавати адаптивне значення швидкості росту плідників і величини плодючості. Адже логічним є те, що збільшення швидкості росту самок і разом з нею зростання їх плодючості є адаптивною відповіддю на покращення умов існування популяцій, які повинні вести до збільшення чисельності, а відповідно і до поліпшення якості статевих продуктів і зростання життєстійкості нащадків. Проте, як показують експериментальні дослідження, у багатьох видів риб із збільшенням росту і плодючості зменшується і величина, і маса, і жирність ікринок, а разом з цим - і тривалість життя личинок за голодування. Тобто, якщо ці якості ікринок дійсно мають велике значення для виживання нащадків, то у даному випадку їх зміни повинні вести не до збільшення, а до зменшення їх чисельності. В дійсності ж із зростанням швидкості росту плідників і їх плодючості відбувається одночасно і покращення якості статевих продуктів і збільшення життєстійкості нащадків.
Це пояснюється багатьма факторами і перш за все, необхідно відмітити, що фізіологічні властивості плідників передаються нащадкам. Встановлено, що самки осетра, форелі, коропа, ляща, тарані, маринки та інших, які характеризуються кращим обміном, демонстрували більшу життєздатність і виживаність. Разом з тим виявилось, що інтенсивність обміну може зростати із збільшенням приросту тіла. Адже на коропі показано, що швидкість росту самиць також передається нащадкам. Багаторічний відбір кращої молоді райдужної форелі від кращих за ростом, плодючістю і якістю статевих продуктів плідників привів до суттєвого зростання швидкості росту риб і прискорення вистигання, збільшення плодючості і виживаності.
Розглянемо більш детально питання щодо впливу швидкості росту плідників риб на виживаність їх нащадків на ранніх етапах постембріонального розвитку. З цього приводу існує досить широка інформація, отримана на багатьох видах риб. Ікра від тугорослої самки ляща виявилась більш чутливою до високої температури в період інкубації і дала більш високий відхід, ніж ікра, отримана від самиць з гарним ростом. Спеціальними дослідами на тарані встановлено, що із зростанням приросту самиць виживаність личинок, які живились, збільшувалась, а тривалість життя голодуючих, навпаки - зменшилась, що пов’язано з кращим обміном речовин.
Самиці судака віком від 3 до 5 років, які мали більший розмір, дали зародків з кращою виживаністю і з більшою інтенсивністю росту, ніж тугорослі самиці того ж віку.
Узагальнюючи ці та інші багаточисельні дані, вчені дійшли висновку про те, що виживаність мальків, які народились від швидкоростучих самок, вище, ніж від тугорослих самиць.
Важливим є також питання щодо існування взаємозв’язку між швидкістю росту плідників і чисельністю їх нащадків. У цілому виявилось, що плідники, які характеризувались більш високим ростом, давали загалом і більше нащадків.