ТЕОРЕТИЧНІ ОСНОВИ РИБНИЦТВА - І.М. Шерман - 2011
3. ТЕОРЕТИЧНІ ОСНОВИ ВЕДЕННЯ РИБНИЧИХ ТЕХНОЛОГІЧНИХ ПРОЦЕСІВ
3.2 Основи ефективного відтворення риб
Формування ремонтно-маточних стад - це безумовна вагома базова складова ведення рибничих технологічних процесів, яка у свою чергу передбачає володіння основами ефективного відтворення риб.
Світова іхтіофауна, на думку різних авторів, включає від 16 до 22 тисяч видів риб, що зумовлює їх величезну різноманітність, зумовлену боротьбою за виживання протягом тривалого філогенезу. Адаптація видів до умов середовища існування протягом своєї історії здійснюється в різних напрямках, що привело до екологічного диференціювання риб, в основі якого закладений принцип поділу згідно місць їх існування.
В зв'язку з цим Г.В. Нікольський пише, що види риб, за аналогією з іншими живими організмами, населяють властиві їм місця існування. Виходячи з пропонованої концепції місцеіснування риб, в значній мірі характеризує вид в плані його взаємовідносин з екологічними параметрами
середовища і суттєвим чином визначає біологію розмноження. Відповідно за рівнем пристосування до конкретного місцеіснування певною мірою умовно розрізняють такі екологічні групи: морські, прісноводні, прохідні і солонуватоводні.
В зв'язку з тим, що знання про екологічні групи риб мають велике значення при розгляданні питань, що пов'язані з природним відтворенням, доцільно розглядати ці відомості перед конкретним переходом до суті, яка орієнтована на вивчення ефективного відтворення і як наслідок розведення риб.
Морські риби. Живуть в солоній морській воді протягом всього свого життя. Серед цієї екологічної групи виділяють пелагічних риб, що населяють товщу води, і донних, які живуть біля дна, в придонних шарах води. Морських риб також підрозділяють на океанічних, що живуть у відкритих частинах і поверхневих шарах океану, також неретичних, що населяють прибережні морські води, а також глибоководних, або аббісальних. Останні в свою чергу підрозділяють на батіпелагічних і донних.
Прісноводні риби. Риби, що живуть в прісній воді протягом всього свого життя, як правило, в солонуватій воді не зустрічаються. Прісноводні риби поділяються на реофільних, пристосованих до життя в проточній воді, і лімнофільних, пристосованих до життя в стоячій воді. В свою чергу реофільні і лімнофільні підрозділяються на пелагічних (що населяють товщу води), придонних і донних.
Прохідні риби. Риби, що проводять частину свого життя в прісній воді, а частину — в солоній. Процес розмноження їх пов'язаний з переходом із морської солоної води в прісну річкову, або навпаки із прісної в солону. В залежності від місць, де вони живляться, їх підрозділяють на трофічно морських і трофічно прісноводних.
Солонуватоводні риби. Риби, які населяють опріснені ділянки морів, естуарій і внутрішні моря, які характеризуються зниженою солоністю, завдячуючи річковим системам, які несуть прісні води. Підрозділяють на напівпрохідних, які заходять для розмноження в пониззях рік, а живляться в солонуватій воді, і власне солонуватоводних, які постійно живуть в опріснених водах.
Характерною особливістю абсолютної більшості представників розглянутих груп є те, що за досягнення статевої зрілості вони здійснюють більш-менш виражені нерестові міграції, в процесі яких завершується статевий розвиток, проходить достигання статевих клітин в гонадах, що передує нересту.
Морські риби постійно живуть і розмножуються в морі. Для прісноводних риб, що проживають в ріках і прісних озерах, характерна слабка вираженість міграцій, вони нерестять безпосередньо біля місць постійного існування. Прохідні риби живуть в абсолютній більшості в морі, а на нерест заходять в ріки, по яких багато з них проходять сотні кілометрів, долаючи течію, водопади і пороги. Напівпрохідні риби локалізуються в опріснених ділянках моря, а для розмноження заходять в ріки, не піднімаючись по них так далеко, як прохідні.
В переднерестовий період яєчники самок несуть в собі статеві продукти, що відповідають завершеній IV стадії стиглості. В процесі підходу до місць нересту, під дією гонадотропного гормону гіпофізу вони зазнають змін, завершується процес овогенезу і проходить овуляція.
Процес овуляції досить складний, для забезпечення нормального його протікання необхідна наявність комплексу біотичних і абіотичних факторів, параметри яких повинні відповідати видоспецифічним особливостям риб. Серед екологічних факторів, необхідних для забезпечення процесу овуляції, виключне значення має температура води, хімічний режим, наявність або відсутність течії, присутність самців, для багатьох видів риб необхідний також специфічний нерестовий субстрат.
В зв'язку з нерестом очевидну цікавість мають дослідження Н.Л. Гербильського, який встановив існування специфічної системи нейрогуморальної регуляції процесу нересту. При цьому, якщо після настання нерестових температур, окремі із названих факторів середовища незадовільні, не відповідають вимогам, овуляція не настає. Такий механізм має для риб пристосувальне значення, яєчники звільняються від овоцитів старшої генерації, і починається новий цикл розвитку.
За нормальних екологічних обставин, що відповідають видоспецифічним особливостям риб, відбувається овуляція, після чого проходить шлюбний акт, який називається нерестом, а місце нересту зветься нерестовищем.
Місця нересту, з урахуванням видоспецифічних особливостей, відповідають вимогам оптимуму для ембріонального і раннього постембріонального періоду життя риб. За аналогією з місцями існування риб, існує диференціювання риб, в основу якого покладений принцип поділу риб по відношенню до нерестових субстратів.
Іншими словами, для відкладання ікри риби вибирають такі ділянки, умови яких відповідають їх біологічним особливостям в період осіменіння, запліднення і розвитку ембріона, раннього постембріогенезу, що слід розглядати в плані адаптації виду в процесі філогенезу.
Г.В. Нікольський, в залежності від особливостей умов розмноження, розвитку і, в першу чергу, того місцеіснування, де проходить нерест, виділяє наступні екологічні групи риб: літофіли, фітофіли, псамофіли, пелагофіли, остракофіли.
Літофіли. Розмножуються, відкладаючи ікру на кам'янистих субстратах, зазвичай в ріках, на течії або на дні оліготрофних озер і прибережних ділянках морів, де, як правило, але не завжди, є сприятливі умови для дихання. До цієї групи відносяться: осетер, білуга, севрюга, стерлядь, сьомга, кета, чавича, горбуша, рибець.
Фітофіли. Розмножуються, відкладаючи ікру на рослинні субстрати за малої проточності або в стоячій воді на вегетуючі або відмерлі рослини. При цьому умови дихання дуже сильно варіюються. До цієї групи відносяться: лящ, сазан, щука, окунь, плітка, карась, сом.
Псамофіли. Розмножуються, відкладаючи ікру на пісок, іноді прикріплюють її до підмитих коренів рослин. Оболонки ікринок часто інкрустуються піщинками, розвиваються зазвичай в сприятливих умовах дихання. До цієї групи можуть бути віднесені: піскарі і деякі види гольців.
Пелагофіли. Розмножуються, викидаючи ікру в товщу води. Ікра і вільні ембріони знаходяться в товщі води в завислому стані, вільно перемішуються під дією течій, хвиль і вітрових явищ. Розвиток, як правило, проходить в сприятливих для дихання умовах. До цієї групи можуть бути віднесені: багато видів оселедців, тріскових, камбал, чехонь, білий і чорний амури, білий і строкатий товстолобики.
Остракофіли. Розмножуються, відкладаючи ікру в мантійну або зяброву порожнину молюсків, іноді під панцир крабів і інших тварин. Ікра зазвичай розвивається в не досить задовільних умовах дихання. До цієї групи відносяться гірчаки.
Очевидно, що розглянуті групи риб не охоплюють всієї різноманітності іхтіофауни. Поряд з наведеними екологічними групами і видами, що в них входять, існують і живонароджуючі риби, у яких ембріогенез повністю або частково відбувається всередині материнського організму. В складі іхтіофауни є багато видів, які за характером розмноження займають проміжне положення, відносячись до різних груп. В залежності від умов середовища рибець, кутум, в'язь можуть відкладати ікру на кам'яному ґрунті або на рослинності, а судак і корюшка на пісок або рослинність.
Знання екологічних груп в зв'язку з особливостями розмноження в ланцюгу проблем, пов'язаних з розведенням риб, має виключну значимість, що обумовлено необхідністю обґрунтованого біологічного підходу до технології риборозведення.
В іхтіології взагалі і в риборозведенні, зокрема, існує поняття «нерестова температура». Це зумовлено тим, що за наявності суми факторів, необхідних для нересту риб, температура води виступає сигнальним фактором, що запускає механізм нересту. Проте для кожного виду риб потрібен точно визначений діапазон температур води, за якого можливий нерест, що повинно поєднуватися з наявністю відповідних нерестовищ. При цьому тривалість нересту, або нерестової кампанії, тісно зв'язана з динамікою температури води. Звичайно, спостерігаючи нерестову кампанію, спеціаліст керується параметрами коливання або діапазону температур, температурними межами, у яких конкретний вид починає нерест, або його закінчує. Звідси очевидно, що тривалість нересту або нерестової кампанії залежить від того, з якою швидкістю буде змінюватись температура в процесі нересту, або скільки потрібно діб, щоб температура води змінилась до величини температури води, початку нересту, до температури води, відповідної до закінчення нересту. В умовах штучного відтворення, регулюючи температуру води, можливо безпосередньо впливати на тривалість окремих технологічних циклів, виходячи з особливостей певних підприємств.
Розглянуте положення має загальний характер і пояснює взаємозв'язок між тривалістю нересту і температурою води. В залежності від динаміки температурного режиму тривалість нересту, або нерестової компанії може різко скорочуватись або суттєво розтягуватись. Поряд з цим на тривалість нересту має безпосередній вплив тип відкладання ікри. За порційного відкладання ікри характерний більш тривалий період нересту, ніж для риб з одночасним ікровикиданням. Досить значна також роль вікової структури стада; характерно чим довший віковий ряд особин, що представляють нерестову популяцію, тим триваліший період нересту. Коли в складі нерестової популяції є певні екологічні групи, то ця особливість теж може бути причиною розтягнутості нересту. За інших рівних умов, різкі зміни температури води, що виходять за межі нерестової, негативно впливають на результативність нересту. Фіналом і логічним завершенням нересту стає контакт ікри зі спермою, її запліднення і утворення зиготи.
Наведені екологічні особливості розмноження потрібно враховувати при створенні та адаптації технологій з риборозведення, маючи достатню спеціальну підготовку з цих питань.
Для іхтіофауни помірних та високих широт, де відносно чітко спостерігається сезонність, що обумовлює закономірну зміну температурного режиму водойм, об'єктивна наявність двох біологічних груп риб. Загальновідомо, що час нересту різних видів риб досить різноманітний, але пристосований до відповідних сезонів року, що дозволяє, виходячи з реалій, виділити наступні групи риб:
- Весняно - літньонерестуючі, що нерестують у весняно-літній період, (щука, окунь, судак, тараня, лящ, сазан, осетер, білуга, рибець, севрюга);
- Осінньо - зимовонерестуючі, що нерестують в осінньо-зимовий період ( сьомга, кета, чавича, кижуч, форель, миньок, сиги).
Для риб, що нерестують у весняно-літній період, характерна протяжність нересту з березня до серпня, а для осінньо-зимово нерестуючих - з вересня до січня. При цьому риби одного виду і навіть однієї популяції можуть мати різні календарні терміни ікровикидання.
Неодноразово відмічено, що для окремих груп прохідних риб, які відносяться до одного виду, характерні різні терміни заходу в ріки на нерест. В зв'язку з цим Л.С.Берг дійшов до висновку, що деякі лососеві і осетрові риби об'єктивно мають озимі і ярі раси. Ярі особини здійснюють нерестові міграції із моря в ріки, де вони відкладають ікру цього ж року, і повертаються на нагул в море. Озимі особини здійснюють нерестові міграції із моря в ріки, де зимують, а нерестують вже в наступному році, після чого повертаються в море на нагул. При цьому між ярими і озимими формами існує диференціація місць нересту. Місця нерестовищ озимих риб розташовані значно вище по течії рік, ніж в ярових. Така своєрідна спеціалізація всередині виду по відношенню до розміщення місць нересту має пристосувальний характер, що забезпечує певну стабільність виду в ареалі. Вид отримує можливість використання нерестовищ різних ділянок ріки на значних відстанях, що підвищує ефективність відтворення і збільшує можливості збереження і розквіту виду, дозволяє уникнути можливих несприятливих умов, які періодично виникають на окремих нерестовищах, сприяє кращому виживанню нащадків.
Різні біологічні групи всередині популяції є наслідком особливостей ареалу виду, але характерним є особливість певної мінливості, коли
відповідні умови відсутні, внутрішньопопуляційна диференціація не проявляється.
Диференціацію в межах популяції належить розглядати в якості пристосувального механізму до умов розмноження і розвитку. Внутрішньо- популяційна диференціація — не що інше, як адаптація до умов ареалу, що дозволяє найповніше використати його можливості. Інформація про диференціацію складу популяцій становить не тільки теоретичну зацікавленість, відповідні знання в цій галузі дозволяють кваліфіковано вирішувати багато практичних завдань, пов'язаних з лімітуванням вилову цінних прохідних риб, розміщенням риборозвідних заводів, свідомо планувати технологією риборозведення. Виключне значення за штучного риборозведення набувають роботи з різними біологічними групами одного виду при необхідному кількісному співвідношенні. Тільки таким чином можна зберегти генетичну і екологічну структуру популяції і забезпечити ріст чисельності, що досить вагомо за штучного відтворення осетрових і лососевих.
Досить важливим моментом, що визначає належність особини до статі, є характер поведінки самок і самців. В елементарному випадку репродуктивна поведінка пов'язана з утворенням нерестових скупчень і спільним викидом статевих продуктів на нерестовищах. Більш складна поведінка обумовлена «шлюбною грою», переслідуванням самцями самок (короп, щука). Іще більш різноманітна поведінка пов'язана з підготовкою самцями нерестовища, побудовою своєрідного «гнізда» і приваблюванням самки (тиляпії). Під час репродуктивного періоду самці проявляють агресивність, що обумовлено проявами демонстративної поведінки, залицяння, охорони території.
Після викиду статевих продуктів для багатьох видів характерна та чи інша форма турботи про нащадків: обмахування ікри в процесі ембріогенезу, виношуванні ікри в ротовій порожнині, догляд за нащадками в період раннього постембріогенезу. Така поведінка самців і самок є результатом включення природжених стереотипів під дією гормональної системи, однією з ланок якої є самі статеві залози.
У VIII столітті вчені фактично не мали чіткої уяви про те, як розмножуються тварини, що населяють водні об’єкти. Навіть найвидатніші натуралісти того часу помилялись відносно ролі сперматозоїдів в процесі запліднення яйцеклітин, вважали їх сторонньою, несуттєво - важливою частиною статевих клітин самців.
Особливості ембріогенезу і ранній постембріогенез є складовими процесу відтворення риб в природних умовах і розведення. Важко назвати ланки технологічного циклу розведення риб, які були б більш-менш значими для процесу взагалі. Всі ланки взаємопов'язані, послідовні і взаємозалежні. Втрати, понесені на будь-якому етапі технологічного циклу, не можуть бути надолужені наступними зусиллями оптимізації технології риборозведення. Проте загальновідомо, що максимальні втрати при риборозведенні спостерігаються в період ембріогенезу і раннього постембріогенезу.
Після запліднення ікринки і утворення зиготи відразу починається процес розвитку ембріона. Тривалість ембріогенезу у риб варіює дуже в широкому діапазоні від десятків годин до декількох місяців, що визначається видоспецифічними особливостями різних систематичних груп. Тривалість ембріонального розвитку може зазнавати суттєвих змін і в межах одного виду, що, за інших рівних факторах, залежить від температури води. Тим часом для цілого ряду видів риб установлено, що температура води в межах норми впливає на тривалість ембріогенезу, але при цьому число градусо-днів характеризується практично постійними величинами. Так, для форелі це число складає 410 і фактично не залежить від температури води, на фоні якої можливий нормальний ембріональний розвиток:
Температура,°С |
Тривалість ембріогенезу, діб |
Число градусо-днів |
2 |
205 |
2 х 205 = 410 |
5 |
82 |
5 х 82 = 410 |
10 |
41 |
10 х 41 = 410 |
Із цього прикладу очевидно, що, змінюючи температуру води в процесі ембріогенезу, можна впливати на тривалість ембріогенезу. Це положення має закономірний характер і правильне для умов, близьких до вимог, що ставляться видом до термічного режиму в процесі ембріогенезу.
При цьому необхідно враховувати той факт, що кожний вид риб характеризується певним діапазоном нерестових температур, який може коливатись у вузьких (стенобіонти) або широких (еврибіонти) межах. В усякому випадку, що виключно важливо для розведення риб, існують оптимальні нерестові температури.
В зв'язку з цим, розглядаючи можливості терморегуляції в умовах заводського відтворювання певних видів риб, необхідно враховувати таке поняття, як оптимальний температурний режим, його межі, видоспецифічні вимоги об'єкту розведення.
Відомо, що температура замерзання морської води, через розчинені в ній солі, нижче нуля, що не заважає нормально розвиватись ікринкам ряду видів морських риб за мінусових температур. Зміна тривалості інкубаційного періоду за коливання температури на один градус при низьких температурах набагато значніша, ніж при високих. Проте ці зміни в ряді випадків не мають пропорційного характеру.
В процесі ембріогенезу на ікру, що розвивається, досить суттєво впливає світло. В ряді випадків збільшення освітленості справляє стимулюючий вплив, прискорюючи розвиток ікри. Так, ікра камбали в тіні, за однакових температур, розвивається на півтори-дві доби довше, ніж на світлі. Прискорення розвитку ікри на світлі і уповільнення в темряві відмічено у севрюги.
Діаметрально протилежно реагує на світло ікра риб, ембріогенез яких в природних умовах проходить в темряві. Так, форель відкладає ікру на нерестовищах в грунт, представлений гравійно - гальковими субстратами, а ембріогенез відбувається в місцях, які недоступні для сонячних променів, при освітленні спостерігається збільшення інкубаційного періоду на чотири-п'ять діб.
За інтенсивного освітлення ікри і вільних ембріонів у лососів спостерігається порушення нормального обміну речовин, що супроводжується масовою загибеллю. В зв'язку з такою видоспецифічністю в процесі онтогенезу у личинок цих риб виробилась негативна реакція на світло, або негативний фототаксис, вони активно ховаються від світла, локалізуються в затемнених місцях. Не виключено, що активний відхід із освітленої зони, поряд з іншими причинами, є і однією з форм захисту від ворогів.
Зміна параметрів середовища в межах амплітуди, відповідної адаптаційним можливостям виду, викликає у ембріона, що розвивається, певні морфологічні зміни, що мають адаптаційний характер. Проведені дослідження показали, що зниження температури води або підвищення її мінералізації в період сегментації ембріона приводить до збільшення кількості хребців у хвостовому відділі, що можна розглядати в якості специфічної реакції пристосувального характеру, обумовленої змінами щільності води. Підвищення температури води і зниження її мінералізації дає протилежний ефект, кількість сегментів в хвостовому відділі, а отже і кількість хребців у дорослих особин зменшується, для ряду видів оселедців, лососів.
В процесі ембріогенезу міцність оболонок ікринок зазнає суттєвих змін, що чітко простежується на ікрі осетрових. В осетрових міцність оболонки не тільки підлягає змінам в процесі ембріогенезу, але і є специфічною для кожного виду. Найбільш міцною є ікра білуги, розвиток якої відбувається на ділянках ріки з досить сильною течією. При цьому нерестовища білуги розташовані найбільш високо по течії. Поступає їй в міцності лише ікра нерестуючого нижче осетра, мінімальна міцність характерна для ікри севрюги, нерестовища якої розташовані найнижче по течії.
У риб, які живуть в водоймах, що пересихають, або які відкладають ікру у верхній частині припливно-відпливної зони, під час обсихання ікри розвиток може затримуватись, настає діапауза, тривалість якої може досягати трьох місяців, а після контакту з водою ембріогенез продовжиться.
Викльовування ембріона з оболонки відбувається завдяки зниженню її міцності, що досягається дією спеціального ферменту, який виділяється залозами вилуплювання, розміщеними на головній частині ембріона. При механічному розриві оболонки фермент вилуплювання не виробляється. Подальший розвиток вільного ембріону після викльову із ікринки, як і до викльову, являє собою видоспецифічний процес, що має пристосувальний характер.
У зв'язку з цим необхідно звернути увагу на теорію етапності розвитку риб, що поліпшить розуміння і сприйняття тих змін, які характерні для ембріогенезу і раннього постембріогенезу. Докладний аналіз морфології і фізіології розмноження на фоні екології дозволив В.В.Васнецову (по Г.В.Нікольському) встановити, що протягом життя особини змінюються її біологічні особливості і взаємодія з середовищем, що і покладено в теорію етапності розвитку. У зв'язку з розглядаємою теорією стосовно до ембріогенезу і раннього постембріогенезу доцільно підкреслити, що кожний етап характерний для всієї різноманітності світової іхтіофауни, але специфіка умов розвитку представників кожної екологічної групи і видоспецифічні особливості привносять певну індивідуальність, обумовлену особливостями будови і фізіології.
Виходячи з викладеного, доцільно окреслити коло питань, що підлягають вивченню. Поняття ембріогенез в спеціальній літературі не несе тлумачення, що дозволило викладати матеріал, не зупиняючись на часових рамках. Практично всі класичні праці і окремі публікації прикладного характеру пропонують розуміти під ембріогенезом або ембріональним періодом розвитку часовий інтервал — від моменту запліднення ікри до переходу передличинки або вільного ембріона на активне живлення, що свідчить про завершеність ембріогенезу, а особина досягла личинкової стадії розвитку. Відносно ж значення поняття постембріогенез і його часові рамки існують різні думки. Окремі спеціальні дослідження обмежується раннім постембріогенезом, що дозволяє акцентувати увагу на тому, що не виходить за межі поняття постембріогенез. Одне із широко розповсюджених формулювань пропонує вважати раннім постембріогенезом період розвитку риби від початку активного живлення до формування особини з усіма характерними ознаками, притаманними виду, іншими словами досягнення малькової стадії розвитку. Таким чином, очевидно, що відрізок часу між личинкою і мальком може розглядатись в якості раннього постембріогенезу, що є частиною загального постембріогенезу, охоплюючого часовий інтервал між личинкою і всією подальшою тривалістю життя риб.
Повертаючись до раннього постембріогенезу, переконуємося, що вільні ембріони, які виклюнулись з ікринки, в залежності від особливостей виду і температурного режиму в процесі розвитку, за інших сприятливих факторів середовища, можуть більш-менш тривалий час живитись за рахунок жовткового мішка.
Для риб з весняно-літнім нерестом, відносно коротким періодом ембріогенезу процес так званого жовткового живлення не тривалий. Тривале жовткове живлення характерне для риб з осінньо-зимовим нерестом, відносно довгим ембріогенезом. Можливість нормального перебігу ембріогенезу і раннього постембріогенезу як у передличинок, так і у личинок, обумовлена наявністю достатньої кількості розчиненого у воді кисню, і нормальним розвитком відповідних органів дихання. У вільних ембріонів і личинок різних систематичних груп риб пристосування до дихання досить різноманітне. В процесі онтогенезу органи дихання риб зазнають суттєвих змін, що пов'язано з певними змінами в способі життя. У ембріонів і личинок риб до розвитку дефінітивних зябрів засвоєння кисню здійснюється через систему кровоносних судин на жовтковому мішку і плавцевій складці. При цьому відмічено, що за сприятливого кисневого режиму в ембріонів і личинок кровоносна дихальна система розвинена слабше, а за пониження вмісту кисню у воді спостерігається більш сильний розвиток капілярної сітки, що сприяє поліпшенню забезпечення організму киснем.
В процесі подальшого розвитку, що супроводжується розсмоктуванням жовткового мішка, скорочується і дихальна кровоносна сітка, але це явище компенсується за рахунок збільшення кровоносної сітки в плавцевих складках.
Личинки дводишних риб до розвитку дефінітивних органів дихання, властивих дорослим особинам, мають личинкові (зовнішні) зябра, зустрічаються і несправжні зябра або псевдобранхія. В якості ембріонального органу дихання виступають анальні відростки, що характеризуються добрим розвитком кровоносних судин.
Розглядаючи особливості дихальних органів передличинок і личинок, легко помітити наявність певного зв'язку з місцями нересту. Менший розвиток отримали личинкові органи дихання у пелагофільних риб, розвиток яких, як правило, проходить в сприятливих умовах на тлі високого вмісту кисню, розчиненого у воді. Личинкові органи дихання фітофілів досягають високої досконалості і представлені сіткою кровоносних судин не тільки на жовтковому мішку, але і в плавникових складках і зябрових кришках. Проте найбільшого розвитку і досконалості досягають личинкові органи дихання в літофільних риб. Поява формених елементів крові, як переносників кисню і необхідної умови дихання, спостерігається раніше у видів, що розвиваються в менш сприятливих умовах (фітофіли, літофіли, остракофіли), і відбувається звичайно до виходу з оболонки. У ембріонів, що розвиваються в сприятливих кисневих умовах (пелагофіли), формені елементи крові з'являються значно пізніше.
Виклюнувшись із ікринки, передличинка або вільний ембріон має спеціальні функціональні системи, які забезпечують можливості утримання в місцях, які найбільш сприятливі для раннього постембріогенезу. В цей період передличинки і личинки, які ведуть пелагічний спосіб життя, мають своєрідні вирости тіла, які дозволяють їм протистояти течіям і утримуватися в товщі води.
Передличинки фітофільних риб, ікра яких розвивається в прикріпленому стані на рослинах в узбережній зоні, часто мають «цементний» орган, який дозволяє їм фіксуватись на субстраті і утримуватись в поверхневих шарах води з оптимальним кисневим режимом.
Для остракофілів це явище не характерне, таких видів відносно мало, зокрема гірчак, у якого утворюються спеціальні вирости жовткового мішка, якими передличинка утримується в зябрових пелюстках молюска. У псамофілів в процесі раннього постембріогенезу звичайно утримування здійснюється за рахунок збільшення грудних плавців, а в деяких піскарів утворюється подоба присоски.
Деякі особливості ембріогенезу і раннього постембріогенезу пов'язані з живонародженням, що вимагає короткої інформації з цього питання. Морська бельдюга характеризується справжнім живородінням, жовток у ембріонів бельдюги дуже малий, поживне середовище для розвитку ембріона — ембріотроф, формується в порожнині гонад із відмерлих яйцеклітин і ембріонів. Циматогастер із сімейства ембіотоцид характеризується внутрішньоутробним розвитком настільки тривалим, що самці народжуються статевостиглими. У цього виду ембріотроф утворюється із залишків сперми, відмираючих овоцитів і ембріонів. Всмоктування поживних речовин відбувається через поверхню жовткового міхура, а дихання — через дуже розвинені промені плавців. Досить досконалий процес внутрішньоутробного розвитку і в таких живородящих риб як пецилій, гуппі, молінезій і інших акваріумних риб. Для ембріонів цих риб характерний інтенсивний розвиток зовнішньої кровоносної сітки, що забезпечує їм дихальний процес, а оточуючі ембріон фолікули утворюють живильну псевдоплаценту. Високого рівня внутрішньоутробного розвитку досягли ембріони ряду акулових, які народжуються повністю сформованими, а їх довжина може перевищувати 20 см. Ембріогенез у них відбувається в своєрідних матках, які забезпечені живильними утворами на зразок плаценти ссавців.
Видова різноманітність світової іхтіофауни, своєрідність і широка варіабельність кількісних і якісних параметрів середовища проживання
зумовили в процесі адаптації видів до умов існування відмінності в особливостях і характері ембріогенезу окремих систематичних груп.
В зв'язку з тим, що об'єктами розведення є види, які характеризуються ембріональним і раннім постембріональним розвитком, який проходить в зовнішньому середовищі, а не всередині материнського організму, оптимізуючи умови середовища, ми отримуємо реальну можливість управління процесами ембріогенезу і раннього постембріогенезу в умовах риборозведення.
В зв'язку з тим, що основним традиційним і достатньо вивченим об'єктом тепловодного ставового рибництва є короп і його розведенню приділяється особлива увага, необхідно розглянути його ембріогенез і ранній постембріогенез більш детально.
Короп є типовим фітофілом і відкладає ікру на рослинність в стоячій або слаботекучій воді за температури від 17°С і вище, що характерно і для його вихідної форми — сазана, існує інформація, що ефективний нерест є можливим за значно нижчих температур води.
Ікра звичайно жовтого кольору, але зустрічається із зеленуватим відтінком і безбарвна, середній діаметр ікринок 1,5-1,8 мм з невеликим перивітеліновим простором. За кількістю цитоплазми займає одне із перших місць серед ікри риб сімейства коропових. Діаметр жовткового міхура в середньому 1,2 мм. Оболонка ікринок клейка. Тривалість розвитку ікри коропа до виходу із оболонок вільних ембріонів або передличинок залежить в першу чергу від температурних умов. Встановлено, що для нормального розвитку ікри до викльову необхідна певна кількість тепла, для коропа це 60-80 градусо-годин.
Тривалість розвитку ікри коропа в залежності від температури (за даними Ф.М.Суховерхова і А.П.Сіверцева):
Температура води, °С |
22 |
20 |
19 |
17 |
нижче 16 |
Тривалість інкубації, дні |
2,5-3 |
3,5-4 |
4,5-5 |
7-7,5 |
більше 8 |
Сучасні уявлення свідчать про те, що ембріональний період розвитку коропа складається із декількох етапів (рис. 3.1 ).
На І етапі (1, рис.) відбувається утворення перивітелінового простору і бластодиску. У незаплідненій ікринці оболонка щільно прилягає до жовтка. Початком першого етапу онтогенезу є утворення зиготи. Етап продовжується до початку дробіння. Через декілька хвилин після запліднення в ікрі, що знаходиться у воді, відбуваються зміни, пов'язані з проникненням води в ікринку. Це призводить до відшарування оболонки від жовтка і утворення перивітелінового простору. Процес набрякання ікри за температури 19°С триває приблизно годину.

10
Рис. 3.1 Етапи ембріонального розвитку коропа (пояснення за текстом)
Діаметр ікринок збільшується в середньому на одну третину. Одночасно в період набрякання утворюється зародковий диск, або бластодиск.
Активація ікринок, викликана заплідненням, призводить до глибоких змін обміну речовин. Протягом першої години після запліднення, коли настає різке оводнення ікринок, відносний вміст сухих речовин знижується із 30-32 до 10-12% і приблизно в такій кількості залишається до викльову ембріона. Вміст глікогену — основного джерела енергії в період утворення бластодиска — зменшується в 2 рази, а величина аденозинтрифосфорної кислоти, яка займає центральне місце в енергетичному обміні, знижується майже в 3 рази.
На ІІ етапі (2-3, рис) відбувається дробіння бластодиска від двох бластомерів до бластули, збільшуються кількість клітин і зменшуються їх розміри. Ікринка проходить ряд стадій розвитку. Через три години після запліднення наступає стадія дробіння, з'являється перша боріздка, яка поділяє бластодиск на дві клітини — бластомери, а потім настають стадії чотирьох, восьми бластомерів. Через 6 годин з моменту запліднення настає стадія морули крупних клітин. Далі клітини бласкодиску все більше дробляться. Настає стадія морули дрібних клітин. Між бластодиском і жовтком виникає невелика порожнина або бластоцель і наступає стадія бластули. Бластула — це своєрідне утворення — бластодерма, яка розміщена на анімальному полюсі жовтка.
В цілому процес дробіння супроводжується значними внутрішніми енергетичними затратами. За цей період показник АТФ знижується майже в два рази.
В рибничій практиці на стадіях 4-8 бластомерів другого етапу дають оцінку якості ікри відносно нормальності дроблення. Утворення різномірних, асиметрично розміщених бластомерів свідчить про активний розвиток ікри. Саме на стадіях дроблення від 4-8 бластомерів до ранньої морули визначають відсоток запліднення ікри.
На ІІІ етапі (4-5, рис) відбувається обростання жовтка бластодермою, гаструляція і формування зародка. Гаструляція розпочинається з обростання жовтка багатошаровою бластодермою. Через 8-9 годин половина жовтка охоплюється бластодермою. З'являється зародковий валик, його досить чітко видно на стадії замикання жовткової пробки. На тілі зародка помітний розширений головний відділ. Жовткова пробка замикається. Гаструляція завершується повним обростанням бластодермою всього жовтка.
Під час гаструляції відбувається суттєва структурна перебудова, в результаті якої утворюється три зародкових пелюстки: ектодерма, мезодерма і ентодерма.
Обмін речовин під час гаструляції має свої особливості. В цей період створюються основи органогенезу. Після гаструляції кількість фосфору аденозинтрифосфорної кислоти і небілкового азоту знижується, а кількість загального білка зростає. Гаструляція завжди супроводжується підвищеною загибеллю ікри. Тому облік відходів доцільно проводити не раніше проходження цієї стадії.
На IV етапі (6-7, рис) відбувається диференціація головної і тулубної частин зародка. Через 17-20 годин після запліднення ікри тіло зародка охоплює біля 3/5 окружності жовтка. Починається сегментація тіла. В тулубі утворюються перші два-три соміта. Через 22-24 години формуються очні пухирці і продовжується сегментація тіла. Через 24-28 годин за очними пухирцями в районі довгастого мозку з'являються слухові плакоди. Кількість сомітів досягає 9-11. Очні бокали (зачатки очей) здобувають щілевидні заглибини.
На V етапі (8, рис) відособлюється хвостовий відділ, і зародок починає рухатись. В результаті відособлення хвостового відділу і росту в довжину зачатка кишкової трубки жовток набуває грушовидної форми. Через 35-45 годин в оці чітко видно кришталик. Кількість сомітів продовжує збільшуватись (більше 20). Тіло ембріона здійснює слабкі рухи. У віці чуть більше двох діб спостерігається сегментація хвостового відділу. До цього часу сегментація тіла майже закінчується. В очах з'являється чорний пігмент. Розрізняються відділи головного мозку. В слухових капсулах утворюються отоліти. При відособленні хвостового відділу і пігментації очей відбуваються певні зміни в обміні речовин: показник АТФ знову збільшується до вихідної величини, проте вміст білка і небілкового азоту залишається таким, як і за гаструляції.
На VI етапі (9, рис) у віці 2,5 діб у ембріона з'являються формені елементи в крові. Кількість сегментів в тулубі 24, в хвостовому відділі — 16.
Очі пігментовані. Сформувалась шкіряна зяброва кришка. Г олова пригнута до жовтка. На рилі перед очима з'являються нюхові ямки, а знизу ротова лійка. Позаду очей утворюються чотири зябрових плакоди. На рівні першого міотома розташовуються маленькі грудні плавці. Ембріон активно перевертається в оболонці.
Ця стадія зародку коропа, як і інших риб, найбільше підходить для транспортування в ізотермічних ящиках, де можлива певна терморегуляція — охолодження, що сприяє уповільненню розвитку ембріона.
На VII етапі (10, рис) ембріон вилуплюється з оболонки. Це останній етап ембріонального періоду розвитку. Через три - п'ять діб інкубації ікри за температури 17-22°С починається викльов ембріонів. Ембріони, що виклюнулись, — передличинки мають відносно слабко пігментовані очі і тіло. Пігментні клітини розміщені на голові і вздовж хорди. Жовтковий мішок великий, грушоподібної форми, сильно пігментований. Вільний ембріон має суцільну плавцеву складку, розширену в хвостовій частині. Голова випрямлена і відділена від хвоста, грудні плавці маленькі. Рот нерухомий, в формі ямки, в нижньому положенні. Кишківник має вигляд прямої здавленої трубки без просвіту. Довжина від рила до кінця хорди (без плавникової складки) складає 4-5 мм.
Після виходу ембріону з оболонки відбуваються суттєві зміни в обміні речовин. Якщо основним джерелом енергії зародка є глікоген, то головним в ендогенному живленні передличинки є жир. Його запаси в два рази вищі (2-2,5%) ніж глікогену (0,7-1,2%). Змінюються і інші показники обміну. Вміст білка збільшується до 11.-14%, сухих речовин — до 19-20%, фосфору — до 300-360 мг %.
В цей період вільні ембріони живляться виключно за рахунок жовткового мішка і малорухливі. Як правило, вони висять; прикріпившись, до рослин, на які була відкладена ікра. Для цього у вільних ембріонів коропа є спеціальні органи, які представлені парними залозами, розміщеними нижче і спереду очей. Передличинки зрідка відриваються і знову прикріплюються; це не тільки рятує їх від ворогів, але і сприяє поліпшенню дихання, для цього періоду характерний позитивний фототаксис.
Таким чином, клейка оболонка ікринок, наявність органів прикріплення вільних ембріонів, які дають можливість висіти на рослинах після викльову, відсутність світлобоязні характеризують коропа як фітофільну рибу, пристосовану до розвитку в стоячих або слабопротічних водоймах із зарослим і замуленим дном.
Необхідно звернути увагу на цю дуже важливу обставину, яку необхідно враховувати в рибогосподарській практиці і, особливо, в сучасному рибництві при широкому використанні заводського способу отримання личинок коропа.
Ікра риб в процесі ембріонального розвитку проходить ряд критичних періодів, коли спостерігається підвищена чутливість ембріонів до різних абіотичних факторів середовища (температура, газовий склад води, солоність, механічний вплив). Ця обставина обумовлена тим, що в періоди підвищеної чутливості відбуваються значні зміни, викликані перебудовою обміну речовин зародка, що розвивається, а порушення цих процесів приводить до масової загибелі.
Критичні періоди в розвитку коропа, як і у більшості нерестуючих навесні риб, припадають на стадії від початку дроблення до морули дрібних клітин, гаструляції, безпосередньо перед викльовом і в період виходу ембріона з оболонки. Саме на цих стадіях ембріогенезу, особливо на початку дробіння, вступу ікри в стадію ранньої гаструли і замикання жовткової пробки, перед вилуплюванням і в момент виходу вільного ембріону з оболонки, спостерігається масова загибель. При цьому необхідно відзначити, що після проходження критичного періоду, загибель ембріонів спостерігається не відразу, а через деякий час, частіше перед настанням наступної стадії розвитку.
В критичні періоди необхідно забезпечити оптимальні умови для розвитку ікри: підтримувати в інкубаційних апаратах постійну, дещо підвищену витрату води, не допускати різких (більше 2°С) температурних коливань, оберігати ікру від різних механічних впливів, виключити проведення санітарно-профілактичних заходів.
Личинковий і мальковий періоди розвитку коропа. В ранньому періоді з моменту вилуплення із оболонки короп проходить 9 етапів розвитку, які В.В.Васнецов позначив літерами: А, В, СІ, С2, Dl, D2, Е, F, G.
Етап А — передличинка відноситься до ембріонального періоду розвитку. Етапи В, СІ, С2, Di, D2, Е характеризують личинковий, F, G — мальковий періоди (рис. 8).

Рис. 3.2. Личинковий та мальковий період розвитку коропа
В рибницькій практиці строки пересадки молоді в малькові або вирощувальні стави визначаються не віком, а етапом розвитку. Зарибнення малькових або вирощувальних ставів рекомендується здійснювати личинками на етапі змішаного живлення. При цьому тривалість підрощування в малькових ставах визначається часом, необхідним для завершення личинкового періоду розвитку і досягненні стадії малька.
Тривалість кожного етапу залежить від температури води, забезпеченості кормом, хімічного режиму і селекційних особливостей коропа (табл. 3.9.).
Таблиця 3.9. Тривалість етапів розвитку коропа в залежності від температури води
Етап |
Температура води,°С |
Тривалість розвитку, діб |
А |
23,8 |
1 і більше |
В |
22,6 |
2,0 |
С1 |
24,0 |
2,0 |
С2 |
24,8 |
1,5 |
D1 |
25,5 |
1,5-2,0 |
D2 |
26,0 |
3,0 |
Е |
23,1 |
2,0 |
F |
23,3 |
3,5 |
G |
23,8 |
18-20 |
Знання особливостей ембріогенезу і раннього постембріогенезу коропа дозволяє свідомо займатись його штучним відтворенням в ставових умовах і спеціалізованих інкубаційних цехах.
Ембріональний розвиток рослиноїдних риб. За своєю значимістю рослиноїдні риби, до яких відноситься типовий макрофітофаг — білий амур і типовий фітопланктофаг — білий товстолобик, є основними компонентами полікультури тепловодних ставових господарств, досить значимими в рибництві рік, озер і водосховищ. До цієї групи також відносяться, через схожість штучного відтворення і походження, строкатий товстолобик і чорний амур, які за характером живлення не є класичними рослиноїдними рибами. Строкатий товстолобик живиться зоо- і фітопланктоном, а чорний амур споживає молюсків.
В умовах штучного розведення статеві залози рослиноїдних риб досягають 4 стадії розвитку, подальший розвиток репродуктивної системи припиняється, і для отримання зрілих статевих продуктів необхідне застосування фізіологічних стимуляторів. В зв'язку з цим для опанування технологією штучного розведення рослиноїдних риб необхідно мати чітку уяву про характерні аномалії ембріогенезу і раннього постембріогенезу, встановлених в процесі тривалих спеціальних досліджень. В біології розмноження і розвитку білого і чорного амурів та білого і строкатого товстолобиків багато спільного, тому в таблиці 2 і на рисунку 9.2 подається
опис характеру розвитку рослиноїдних риб в ранній період життя на прикладі білого амура, а відмінності наведені в таблиці 3.10
Таблиця 3.10. Характеристика розвитку білого амура
Етап |
Стадія |
|
Ембріональний період |
||
I Оводнення порожнини яйцевою оболонкою і яйцеклітиною (поява перивіталінового простору)і утворення плазменого горбка— бластодиска |
Стадія 1. Діаметр неоводненої ікринки після запліднення 1,2-1,3 мм. Яйцева оболонка щільно прилягає до поверхні яйця, вона неклейка і представлена первинною радіальною оболонкою. Ікра прозора, безбарвна або злегка жовтувата. Стадія 2. Вік 10 хв. Після запліднення. Відмежування яйцевої оболонки від жовтка і концентрація плазми у вигляді прозорої серповидної зони. Стадія 3. Вік 40 хв. Утворення чітко окресленого бластодиску. Головним чином завершується оводнення перивітелінового простору. Діаметр ікринки 3,8-4 мм. а власне яйця 1,2-1,3 мм. Такий величезний перивітеліновий простір зменшує масу ікринок і забезпечує її плавучість у потоках води; у стоячій воді ікринка спускається на дно. |
|
II Дробіння бластодиску до бластули |
Стадія 4. Вік 1 година. Утворення двох бластомерів. Стадія 5. Вік 1 год. 20 хв. Утворення чотирьох бластомерів. Стадія 6. Вік 1 год. 40 хв. Утворення восьми бластомерів. Стадія 7. Вік 2 години. Утворення шістнадцяти бластомерів. Стадія 8. Вік 2 год. 30 хв. Великоклітинна морула (рання). Стадія 9. Вік 4 год. 50 хв. Мілкоклітинна морула (пізня). Завершення оводнення перивітелінового простору. Діаметр оболонки 4,32-5,32 мм Стадія 10. Вік 6 годин. Бластула |
|
III Гаструляція — утворення зародкових пластин |
Стадія 11. Вік 7 год. 10 хв. Обростання бластодермою поверхні жовтка. Стадія 12. Вік 10 годин. Жовткова пробка. Стадія 13. Вік 12 год. 10 хв. Замикання жовткової пробки. Зачаток тіла набуває вигляду потовщеного валика, розширений головний відділ його починається на анімальному полюсі, і хвостова частина його закінчується на вегетативному полюсі. |
|
IV Органогенез - диференціація зародкових пластів на зачатки головних органів |
Стадія 14. Вік 15 годин. Утворення Очних міхурів, закладка хорди, початок пігментації мезодерми. Закладка мозкових міхурів. Стадія 15. Вік 18 годин. З'явлення очних бокалів і щілевидного заглиблення в зачатках очей, сегментація тіла міотоми. Хорда добре помітна. |
|
V Обособлення хвостового відділу від жовткового мішка, початок активного руху |
Стадії 16—18. Вік 29-32 години. Випрямлення тіла. Початок енергійних коливальних рухів і обертальних поворотів. Поява на голові і в серцевій ділянці залоз вилуплення |
|
VI Вилуплення зародка із оболонки |
Стадія 19. Вік 34 години. Викльов. Довжина - 5.2 мм. В тулубі 29-31 сегмент, у хвості 12-14. Тіло без пігменту облямоване недиференцированою. плавцевою складкою. В очах чорна пігментна пляма. Малорухомі. В природних умовах пасивно зносяться в товщі води. |
|
VII Утворення ембріональної судинної системи, початок кровообігу. |
Стадія 20. Вік 51 година. Довжина — 6.5 мм. Органи дихання - хвостова вена і кюв'єрові протоки, що розташовані на передній частині жовткового мішка. Рух пасивний. Живляться власним жовтком. |
|
VIII Утворення і початок функціонування рухомого зяберно - щелепового апарату |
Стадії 21—22. Вік 76-96 годин. Довжина 7,5 мм. Початок зяберного дихання. Рот рухомий. Очі повністю пігментовані Передличинки більш рухомі. Живлення жовткове. Чорні пігментні клітини - меланофори - з’являються на голові, над кишківником і в хвостовому відділі, на жовтковому мішку. Редукція ембріональних органів дихання. Закладка плавального міхура. |
|
Личинковий період |
||
I Змішане живлення |
Стадії 23-24. Вік 4,5-6 діб. Довжина 7,5 -7,8 мм. Дихання зяберне. Плавальний міхур заповнений повітрям. Личинка активна, споживає їжу, але продовжує також живитись і за рахунок жовткового мішка. Пігментація тіла посилюється. Личинки плавають в товщі води. На цьому етапі пропонується перевозити личинок на далеку відстань або висаджувати їх в ставки. |
|
II Екзогенне живлення |
Стадія 25. Вік 7 діб. Жовтковий мішок повністю резорбований, живиться виключно зовнішньою їжею. Зяберно - щелепний апарат рухомий. Довжина 7,6 мм |
|
III Формування непарних плавців |
Стадія 26. Вік 9 діб. Довжина 8 мм. Утворення променів в нижній лопасті хвостового плавця. До 16 - ти діб плавці мають плавцеві промені |
|
Стадії 27—28. Кінець хорди загнутий доверху. У хвостовому плавці виїмка. Заповнюється повітрям передній відділ плавального міхура. Закладаються черевні плавці |
||
IV Формування парних плавців |
Стадії 29-30. Вік 20-22 діб. Довжина - 14,2 мм. В парних плавцях утворюються промені |
|
Мальковий період |
||
І Закладення луски |
Вік 1 місяць. Довжина 2 см. Вздовж бічної лінії з’являється луска. |
|
II Мальок з лускатим покривом |
Вік 1,5 міс. Довжина 4-5 см. Прианальна складка зникла. Тіло вкрите лускою. Видно отвори каналу бічної лінії. |
|

Рис. 3.3. Ембріональний період розвитку білого амура:
I етап. Обводнення положовткової порожнини. Стадії: 1 - необводнена ікринка, діаметр 1,3 мм; 2 - початок обводнення навколожовткової порожнини, вік 10 хв.; 3 остаточне обводнення ікринки, діаметр 6 мм, вік 40 хв.
II етап. Дроблення бластодиску. Стадії: 4 - два бластомери, вік 1 год.; 5 - чотири бластомери, вік 1 год. 20 хв.; 6 - вісім бластомерів, вік 1 год. 40 хв.; 7 - шістнадцять еластомерів, вік 2 год.: 8 - крупноклітинна морула, вік 2 год. 30 хв.; 9 - дрібноклітинна морула, вік 4 год. 50 хв.; 10 бластула, вік 6 год. 30 хв.
III етап. Гаструляція. Стадії: 11 - початок гаструляції, вік 7 год. 10 хв.; 12 - жовткова пробка, вік 10 год.; 13 - закінчення гаструляції (замикання жовткової пробки), вік 12 год. 10 хв.
IV етап. Овогенез. Стадії: 14 - утворення очних пухирців, початок сегментації мезодерми, вік 15 год.; 15 - утворення очних бокалів, вік 18 год.
V етап. Обособлення хвостового відділу зародка. Стадії: 16 - початок обособлення хвоста, вік 1 доба 5 год.; 17 - випрямлення і початок згинань тіла зародків, вік 1 доба 5 годин; 18 - початок активного обертання зародка, вік 1 доба 8 год.
VI етап. Викльов ембріона з оболонки. Стадія 19 - ембріон, що тільки виклюнувся довжина тіла 5,2 мм, вік 1 доба 10 год.
VII етап. З'явлення розвиненої ембріональної судинної системи. Стадія: 20 - ембріон з розвиненою судинною системою, довжина тіла 6,5 мм, вік 2 доби 3 год.
VIII етап. Поява рухомого зяберно-щелепового апарату. Стадії: 21 - початок рухомого стану зяберно-щелепового апарату, довжина тіла 7,4 мм, вік 3 доби 22 - редукція ембріональних органів дихання, закладка плавального міхура, довжина тіла 7,5 мм, вік 4 доби.
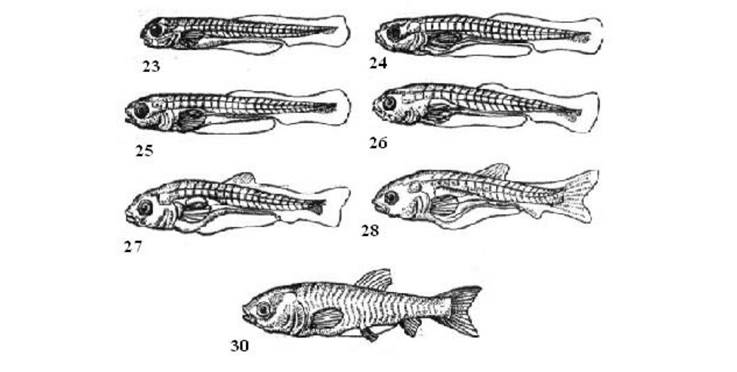
Рис. 3.4. - Личинковий період розвитку білого амура:
І етап. Змішане живлення личинки. Стадії: 23 - початок змішаного живлення личинки з переважанням жовткової їжі, довжина тіла 7,5 мм, вік 4,5 діб; 24 - змішане .живлення личинок з переважанням зовнішньої їжі, довжина тіла 7,8 мм, вік 6 діб.
II етап. Повний перехід н зовнішнє живлення. Стадії: 25 - початок виключно зовнішнього живлення личинки, довжина тіла 7,6 мм, вік 7 діб.
III етап. Утворення непарних плавців. Стадія 26 - утворення скупчення мезенхіми в місцях закладки спинного і анального плавців, довжина тіла 8.1 мм, вік 9 діб; 27 - утворення плавцевих променів в парних плавцях, довжина тіла 9 мм, вік 16 діб.
IV етап. Поява другого відділу плавального міхура. Стадії 28 - поява переднього відділу плавального міхура, закладка черевних плавців, довжина тіла 10,2 мм, вік 18 діб.
V етап. Утворення плавцевих променів в парних плавцях. Стадії 29 - поява плавцевих променів в грудних плавцях, довжина тіла 11,5 мм, вік 20 діб (без рисунка); стадії 30 - наявність променів в грудних і черевних плавцях, довжина тіла 14,2 мм, вік 22 доби.
Таблиця 3.11. - Відмінності в розвитку білого амура, білого і строкатого товстолобиків
Морфологічні і морфометричні ознаки |
Білий амур |
Білий товстолобик |
Строкатий товстолобик |
Ікра (набрякла) |
|||
Середній діаметр, мм оболонки жовткового мішка |
4,38-5,22 1,21-1,36 |
3,80-4,50 1,10-1,20 |
4,82-5,63 1,42-1,50 |
Передличинки, екз |
|||
Кількість міотомів в тулубі в хвості |
29-31 12-14 |
24-26 14-17 |
24-26 14-17 |
Відношення довжини тулуба до довжини хвоста |
2,5 |
2 |
2 |
Наявність чорного пігменту на жовтковому мішку |
Пігмент тільки спереду |
Пігмент спереду і на черевній частині |
Пігмент спереду і слабо на черевній частині |
За штучного розведення риб не завжди вдається забезпечити оптимальні умови для нормального перебігу фізіологічних процесів і правильного розвитку. Це погіршує рибницьку якість ікри, викликає різноманітні порушення в будові зародків і знижує їх життєздатність.
Найчастіше причиною незадовільної якості ікри і аномалій личинок є перетримка ікри в тілі самок після овуляції.
Надійним критерієм якості ікри є відсоток її запліднення і морфологічний малюнок розвитку. Доброякісна ікра має високий відсоток запліднення 90-95%, а іноді і 100%) і не має порушень в розвитку. У недоброякісної часто спостерігається високий відсоток запліднення. При цьому розвиток може відбуватись нормально. Тому лише один показник відсотку запліднення ікри не може правити критерієм якості. Необхідно контролювати процес на різних етапах розвитку, завдяки чому можуть бути виявлені характерні аномалії, що залежать як від якості ікри, так і від умов середовища.
Наприклад, спостерігаються аномалії набрякання, причиною яких є різномірність ікринок, отриманих від однієї самки; така ікра, як правило, добре запліднюється, але має великий відхід в період інкубації і дає значну кількість личинок, для яких характерні різні аномалії розвитку, різні відхилення від норми в будові оболонки ікри, що веде до склеювання ікринок. Це відбувається в тому випадку, якщо ікру в момент заливу води після осіменіння старанно не переміщувати. В місці склейки з ікрою зовнішній шар оболонки розривається і в розрив випинається внутрішній шар.
За аномалії дрібнення бластодиска найбільш частим порушенням є відрив окремих бластомерів і різниця в їх величині (рис. 3.5 ).

Рис. 3.5. - Порушення в розвитку білого товстолобика:
А - відщеплення бластомерів; Б - В - потворний зародок.
За аномалії жовткового мішка жовток у недоброякісній ікрі, порівняно з доброякісною, має більш крупні і неоднорідні гранули. Ембріони з такими змінами жовтка зазвичай доживають до викльову, але з переходом до личинкової стадії розвитку стають нежиттєздатними і масово гинуть.
Несправжній розвиток незаплідненої ікри відбувається своєрідно. Попавши у воду, вона набрякає, на анімальному полюсі утворюється плазматичний горбик, який починає дробитися. Проте дробління виявляється несправжнім, оскільки поділ бластомерів не відбувається до кінця, а утворюються різнорозмірні, асиметрично розміщені псевдобластомери — без'ядерні випуклості плазми (рис. 3.6).

Рис. 3.6. - Зміни незапліднених яєць білого товстолобика:
А - Б - несправжнє дробління; В - Г - несправжнє обростання
Під час такого безладного псевдодробіння незапліднена ікра чітко відрізняється від заплідненої, бластомери якої мають однакові розміри і чіткі контури. Тому найбільш придатними для визначення відсотку запліднення є стадії дробіння від 4-8 бластомерів до ранньої морули. Пізніше межі несправжніх бластомерів зникають, плазма набуває рівну поверхню і починається несправжня гаструляція.
Псевдогаструляція завершується руйнуванням поверхні плазматичного шару і коагуляцією витікаючого з розриву жовтка. Масова загибель незаплідненої ікри співпадає з періодом початку формування тіла зародка в заплідненій ікрі, що потрібно розглядати в якості критерію.
Типовою аномалією розвитку є водянка зародків, яка частіше проявляється на початку формування серця. Вона полягає в надмірному збільшенні і обводненні навколо серцевої порожнини, що в більшості випадків веде до значної деформації серця. Водянка може утворюватись позаду, навколо серцевої порожнини, під переднім або заднім відділом кишківника, в сечовому міхурі, але не виключено утворення такої аномалії і в інших місцях. Життєздатними бувають лише зародки з дуже малозначними ознаками водянки, які в подальшому зникають.
Досить часті і аномалії розвитку за деформації тіла зародку. В цьому випадку характерні викривлення тулуба, хвостового відділу, диспропорцій окремих частин тіла, що звичайно спостерігається і при водянці.
Причиною цих і інших аномалій є безперечна недоброякісність ікри, порушення умов її інкубації, зокрема, за зниженні температури та інших чинників, що потребують уточнень.
Одним із критеріїв оцінки ембріогенезу є діаметр набряклих ікринок. В природному ареалі процес формування фауністичного комплексу, до якого відносяться рослиноїдні риби, здійснюється у водоймах, котрі характеризуються низькою мінералізацією води. Рослиноїдні риби є прісноводними рибами, які характеризуються достатньою пластичністю.
В процесі акліматизації ареал цих видів зазнав суттєвих змін, і рослиноїдні риби, проявивши високу екологічну пластичність,
продемонстрували можливість росту і розвитку в різних екосистемах на тлі досить широкого діапазону мінералізації води. Між тим, переважна більшість напівпрохідних і прохідних видів риб, незважаючи на адаптованість до води з підвищеною мінералізацією, в нерестовий період в якості нерестовища вибирають ділянки річкових систем з прісною водою. В цьому плані рослиноїдні риби, будучи жилими прісноводними рибами, повинні виявляти високу консервативність до підвищеної мінералізації води в процесі ембріогенезу і раннього постембріогенезу. Ця проблема становить певний теоретичний інтерес і виключно значима в практичному плані. Актуальність обумовлена тим, що в ряді регіонів, які успішно освоїли і культивують рослиноїдних риб, спостерігається тенденція не тільки росту мінералізації води вирощувальних і нагульних акваторій, але і акваторій, які використовуються як джерела водопостачання інкубаційних цехів. В зв'язку з цим очевидна необхідність встановлення межі лімітування можливості нормального ембріогенезу за мінералізацією води рослиноїдних риб, що актуально і для інших об'єктів іхтіофауни, яка культивується людиною.
Для отримання об'єктивної інформації були проведені спеціальні дослідження. Інкубація ікри рослиноїдних риб проходила в діапазоні оптимальних температур, активна реакція води була слаболужною-лужною, перманганатна окислюваність відповідала верхнім межам допустимих норм, таблиця 3.12.
Таблиця 3.12. Деякі абіотичні параметри середовища в процесі ембріогенезу
Варіанти |
Середня температура води,°С |
рН |
Перманганатна окиснюваність, м2 О/л |
Мінералізація, г/л |
1 (прісна) |
23,6 |
9,1 |
15,4 |
0,3 |
24,0 |
8,4 |
16,4 |
0,5 |
|
2 (солонувата) |
23,2 |
8,8 |
17,9 |
1,1 |
25,5 |
7,7 |
10,1 |
1,7 |
|
27,7 |
8,1 |
10,0 |
2,7 |
Розвиток ікри рослиноїдних риб, яка інкубується на фоні оптимальних температур, носив типовий характер. За підвищення температури води було відмічено деяке прискорення розвитку. При цьому набрякання ікри білого амура і товстолобиків залежно від мінералізації води відбувається неоднаково. Найбільш інтенсивно цей процес проходить в прісній воді (0,3-0,5 г/л) і продовжується 1 год. 40 хв.-2 год. За збільшення мінералізації води до 1,1-1,7 г/л його тривалість досягає 2 год. 10 хв.-2 год. 30 хв., а при 2,7 г/л - 3 год. 30 хв. (г = 0,96).
Збільшення мінералізації води, яка використовується для водопостачання інкубцехів, не тільки стримує процес набрякання ікринок, але і справляє визначальний вплив на розміри ікри, що набрякла. Встановлено, що між середнім діаметром набряклих ікринок і концентрацією солей існує тісний зворотний зв'язок (г = 0,88), таблиця 3.13.
Таблиця 3.13. Діаметр ікринок рослиноїдних риб, що інкубують за різної мінералізації води
Варіанти |
Мінералізація, г/л |
M±m |
Діаметр, мм |
|
lim |
CV |
|||
1 (прісна) |
0,3 |
4,67±0,06 |
4,30-4,90 |
4,97 |
0,5 |
4,32±0,03 |
4,00-4,50 |
2,62 |
|
1,1 |
3,75+0,04 |
3,30-4,10 |
5,46 |
|
2 (солонувата) |
1,7 |
3,63±0,03 |
2,90-4,15 |
9,51 |
2,7 |
3,09±0,05 |
2,80-3,50 |
7,28 |
|
Про вплив мінералізації води (X, г/л) на розмір ікринок (у1, мм) свідчить відносна помилка апроксимації 3,4%. Зв'язок між цими показниками описується ступеневим рівнянням у1= 3,82 Х-0,17. Зменшення діаметра ікринок за підвищення мінералізації води відбивається на їх загальному об'ємі, що призводить до фактичного зниження навантаження на інкубаційний апарат. Залежність розрахункових норм завантаження (у2, г/л) відносно мінералізації води (X, г/л) описується рівнянням у2= 10,97Х 045, яке має невелику відносну помилку апроксимації — 7,7%.
В зв'язку з цим певну практичну зацікавленість мають норми завантаження інкубаційних апаратів відносно мінералізації води таблиця 3.14.
Таким чином, очевидно, що за різної мінералізації води, та різного набрякання ікринок ембріогенез проходить нормально. При цьому зниження набрякання ікри не потрібно розглядати в плані аномалій. Більш того, за інкубації ікри в солонуватій воді відкриваються можливості більш ефективного використання можливостей інкубаційних цехів, але це потребує підвищення витрат води для ефективного видалення продуктів метаболізму.
Таблиця 3.14. Розрахунок норм завантаження інкубаційних апаратів в залежності від мінералізації води і розмірів ікринок
Варіанти |
Мінералізація води, г/л |
Діаметр ікринок, мм |
Об'єм ікринок, мм3 |
Завантаження в інкубаційний апарат місткістю 100 л |
||
об'єм ікри, л |
маса ікри, г |
кількість, тис. шт |
||||
0,3 |
5,0 |
65,4 |
600 |
450 |
||
1 (прісна) |
0,5 |
4,5 |
50,9 |
775 |
620 |
|
1,1 |
4,1 |
36,1 |
31,4 |
1090 |
870 |
|
2 (солонувата) |
1,7 |
3,9 |
31,0 |
1250 |
1000 |
|
2,7 |
3,3 |
18,8 |
2000 |
1600 |
||
Діаметр набряклих ікринок може розглядатись, як один із критеріїв якості, але не є визначальним. В ряді випадків діаметр ікринок, який не досяг максимуму, після закінчення набрякання може бути більше або менше визначених уявлень. В цьому випадку потрібно проаналізувати абіотичні параметри середовища і не виключати індивідуальні особливості окремих самок, здатних продукувати яйцеклітини більшого або меншого діаметру без зниження їх якісних характеристик, що підтверджується не тільки нормальним ембріогенезом, високим виживанням в ранньому постембріогенезі, але і життєстійкістю і якістю нащадків. Дослідження, виконані з використанням в якості життєздатного матеріалу рослиноїдних риб, ймовірні і для ряду інших видів риб, які є об'єктами штучного риборозведення. Проте, для кожного конкретного виду риб очевидна достатньо висока специфічність.
Періоди і етапи онтогенезу. Проблематика різних аспектів розведення риб в значній мірі обумовлена біологією і екологією індивідуального розвитку. Різноманіття видового складу іхтіофауни, умов існування, видоспецифічних особливостей морфологічного, фізіологічного і біотехнічного характеру виявились об'єктивними передумовами, що визначають термін життя видів, вік досягнення ними статевої зрілості, особливості ембріогенезу і раннього постембріогенезу.
Вищесказане дозволило накопичити значний матеріал, що характеризує онтогенез риб. Проте, тільки В.В.Васнецов, аналізуючи існуючі дані і виконавши великий обсяг спеціальних досліджень, розробив теоретичні основи етапності розвитку риб, сформулювавши головні положення цієї теорії.
Основною концепцією запропонованої ним теорії є те, що незалежно від виду, протягом онтогенезу розвиток риб відбувається не тільки поступово і безперервно, але і переривчасто та стрибкоподібно, наглядно демонструючи філософську категорію переходу кількісних змін в якісні.
В процесі онтогенезу, як вже відмічалось, риб виділяють ембріональний, личинковий, мальковий періоди і період статевої зрілості. Для кожного періоду характерна якісна специфіка. В процесі ембріогенезу відбувається типове ендогенне живлення за рахунок поживних речовин жовткового мішку. Личинки живляться організмами, що живуть в навколишньому середовищі (екзогенне живлення), але в початковий період (від вільного ембріону до личинки) живлення має змішаний характер (ендогенно-екзогенне), і тільки після повного розсмоктування жовткового мішка здійснюється перехід на зовнішнє живлення. Для личинкового періоду життя риб характерна наявність специфічних личинкових органів, які пізніше зникають. З настанням малькового періоду цей процес завершується, личинкові органи повністю зникають, з'являються нові, які роблять малька меншою копією дорослої риби даного виду.
Протягом кожного періоду відбувається низка послідовних етапів, з яких складається певний період. При цьому, кожний етап, в свою чергу, складається зі стадій розвитку, відповідних періоду розвитку. Під періодом розвитку розуміється група послідовних етапів, об'єднаних однаковою якісною специфікою.
Звідси очевидно, що теорія етапності розвитку риб передбачає в процесі онтогенезу чергування повільного, поступового розвитку (кількісні зміни) і перервних, стрибкоподібних (якісних) змін. При цьому спостерігаються різкі зміни у окремих органів і систем. Ці зміни стрімкі і відбуваються протягом декількох годин, а то і хвилин. При цьому необхідно врахувати, що морфологічні зміни найтісніше зв'язані із змінами ряду біологічних особливостей риб, в тому числі на біохімічному і фізіологічному рівнях.
Якісні, або стрибкоподібні зміни в процесі розвитку відбуваються досить рідко, між цим проходить послідовний ріст і повільні, але помітні кількісні зміни. Повільні, іноді латентні зміни, проходять в конкретних межах онтогенезу, що не змінює основної якості характеризуючого даний інтервал розвитку риб. При цьому якісні особливості організму і його відношення із зовнішнім середовищем залишаються незмінними. Інтервали відносної стабільності онтогенезу риб між двома різними якісними змінами іменуються етапами.
Етапи мають послідовний характер, вони носять певні риси і особливості будови організму, що характеризуються специфічними біологічними властивостями і ставлять конкретні умови до середовища існування. Відсутність необхідних умов для нормального перебігу відповідного етапу призводить до зупинки розвитку на певному етапі, що супроводжується пригніченням або повною відсутністю росту і в кінцевому варіанті — загибеллю.
Розглядаючи теорію етапності розвитку риб, ми бачимо, що для онтогенезу характерна як поступова, так і переривчаста динаміка, яка в свою чергу розподіляється на ряд послідовних етапів, протягом яких відбувається збільшення маси і лінійних розмірів, що супроводжується повільними, поступовими змінами, але принципові зміни в будові, фізіології та біології риб виключені. Онтогенез, або розвиток риб — послідовна зміна адаптації організму до середовища існування протягом конкретних етапів розвитку. Подальший розвиток основних положень теорії етапності розвитку риб пов'язаний з дослідженнями С.Г.Крижанівського, який встановив чітку різницю в характері перебігу окремих етапів, визначив їх різний тимчасовий ареал і специфічні особливості в залежності від ряду факторів. Це положення має важливе значення для розробки технології розведення риб. Тому необхідність подальшого вивчення всіх питань в цьому плані відносно видів, що є об'єктами штучного розведення.
Потрібно звернути увагу на те, що в цьому напрямі було виконано дослідження, що дозволило охарактеризувати біологічні і екологічні особливості розвитку риб на різних етапах.
Пізнання особливостей етапів дозволило відпрацювати рекомендації по вирощуванню молоді анадромних риб до скатного стану, перехід до якого характеризується складними і глибокими змінами, сутність яких — підготовка організму до існування в нових умовах. В практичному плані виключне значення має інформація, яка характеризує вплив факторів зовнішнього середовища на організм риб, що знаходяться на різних етапах розвитку.
Вчення про етапність розвитку слід розглядати в якості фундаменту, основи, на якій базується сучасна біотехніка вирощування молоді цінних видів риб і забезпечується ефективність штучного риборозведення.