ОСНОВИ МЕДИЧНОЇ БІОЛОГІЇ - 2012
Взаємодія алельних і неалельних генів. Явище плейотропії. Множинний алелізм. Генетика груп крові
До складу генотипу входить велика кількість генів, які функціонують і взаємодіють як цілісна система. Г. Мендель у своїх дослідах виявив тільки одну форму взаємодії між алельними генами - повне домінування одного алеля і повну рецесивність іншого. Генотип організму не можна розглядати як просту суму незалежних генів, кожен з яких функціонує поза зв’язком з іншими. Фенотипові прояви тієї або іншої ознаки є результатом взаємодії багатьох генів.
Взаємодія генів — результат реалізації генетичної інформації з певної групи генів, які визначають формування, в залежності від їх комбінацій, різних варіантів ознак або фенотипів.
Розрізняють дві основних групи взаємодії генів: взаємодія між алельними генами і взаємодія між неалельними генами. Проте слід розуміти, що це не фізична взаємодія самих генів, а взаємодія первинних і вторинних продуктів, які зумовлять ту чи іншу ознаку.
Взаємодія алельних генів
Гени, які займають ідентичні (гомологічні) локуси в ідентичних (гомологічних) хромосомах, називаються алельними. У кожного організму є лише два алельних гени.
Основні форми взаємодії алельних генів (генів однієї алельної пари): повне
домінування, неповне домінування, наддомінування і кодомінування.
Домінування (домінантність) — переважання в фенотипі гетерозиготного організму одного алеля (домінантного) над іншим (рецесивним) алелем того самого гена. Рецесивність - пригнічення в фенотипі гетерозиготного організму одного алеля (рецесивного) іншим алелем (домінантним) того самого гена. Домінування може бути повним і неповним. У випадку повного домінуваннядомінантна гомозигота (АА) і гетерозигота (Аа) мають однаковий фенотип. Явище повного домінування спостерігалося в дослідах Г. Менделя, де один алельний ген завжди був домінантним, інший - рецесивним. Тому насіння гороху за кольором завжди було або жовтим, або зеленим і не мало іншого кольору. При повному домінуванні в схрещуванні гетерозигот (Аа х Аа,) розщеплення за фенотипом - 3:1, за генотипом - 1:2:1.
За типом повного домінування в людини успадковуються: ямочки на щоках, здатність згортати язик трубочкою, властивість загинати язик назад, вільна долька вуха, а також багато моногенних спадкових хвороб: полідактилія, бульозний епідермоліз, міопатія, кістозно-аденоїдна епітеліома, ахондроплазія, ін.
Неповне домінування - взаємодія алельних генів, коли в гетерозиготного організму домінантний алель не проявляє повністю своєї домінантності, а рецесивний алель того самого гена - своєї рецесивності. При неповному домінуванні фенотип гетерозиготи (Аа) є проміжним між фенотипом домінантної (АА) і рецесивної (аа) гомозигот. Так, у схрещуванні нічної красуні з червоними квітками (АА) і нічної красуні з білими квітками (аа), всі гібриди (Аа) першого покоління F, мали рожеве забарвлення квіток. У схрещуванні гібридів першого покоління F1 між собою (Аа х Аа) у другому поколінні F2 відбувається розщеплення за фенотипом у співвідношенні 1:2:1, яке співпадає із співвідношенням розщеплення за генотипом 1АА:2Аа:1аа, але відхиляється від розщеплення за фенотипом при повному домінуванні (3:1).
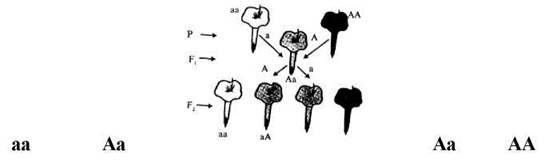
Рис. 19. Моногібридне схрещування "нічної красуні".
За типом неповного домінування в людини успадковуються серпоподібно-клітинна анемія, цистинурія, пильгерова анемія, таласемія, атаксія Фрідрейха, ін. У гомозигот за рецесивним геном цистинурії (аа) в нирках утворюються цистинові камені, у гетерозиггт (Аа) камені не утворюються, а спостерігається лише підвищений вміст цистину в сечі, гомозиготи (АА) - здорові.
Наддомінування - взаємодія алельних генів, при якій домінантний алель у гетерозиготному стані проявляється в фенотипі сильніше, ніж у гомозитному (Аа>АА). Так, у дрозофіли при генотипі АА - нормальна тривалість життя; Аа - подовжена тривалість життя; аа - летальний наслідок.
Кодомінування - взаємодія алельних генів, при якій у фенотипі гетерозиготного організму проявляються обидва алелі того самого гена, в результаті чого утворюється нова ознака. За типом кодомінування в людини успадковується четверта група крові (генотип ІАІВ). У людей цієї групи в еритроцитах крові одночасно наявні антиген А, який контролюється алелем Iа і антиген В - продукт експресії алеля Ів. Алелі Iа і Ів - кодомінантні. Така взаємодія генів спостерігається і при формуванні групи крові MN, гетерозиготи мають на мембранах еритроцитів одночасно 2-а антигени (антиген M і антиген N ).
Взаємодія неалельних генів
Основні форми взаємодії неалельних генів - комплементарність, епістаз і полімерія. Вони переважно видозмінюють класичну формулу розщеплення за фенотипом, встановлену Г. Менделем для дигібридного схрещування (9:3:3:1).
Комплементарність (лат. complementum - доповнення) - форма взаємодії неалельних генів, при якій один ген взаємодоповнює дію іншого, неалельного гена. При одночасній наявності в генотипі комплементарні гени зумовлюють розвиток нової ознаки (новоутворення).
У людини нормальний слух зумовлений комплементарною взаємодією двох домінантних неалельних генів D і Е, з них один визначає розвиток завитки, інший - слухового нерва. Люди з генотипами D-E- мають нормальний слух, з генотипами D-ee і ddE — глухі. У шлюбі, де батьки глухі (DDee х ddEE), всі діти будуть мати нормальний слух (DdEe).
Сіре забарвлення шерсті в мишей контролюється двома генами. Ген А детермінує синтез пігменту, ген В забезпечує скупчення пігменту переважно біля основи і на кінчиках волосся. Схрещування дигетерозигот (АаВbхАаВb) призводить до розщеплення гібридів у співвідношенні 9:3:4 (9 - забарвлення «агуті» : 3 - чорна шерсть : 4 - біла шерсть). Числові співвідношення можуть бути як 9:7; 9:6:1 (видозміна менделівського розщеплення).
Для захисту від вірусів у імунокомпетентних клітинах людини виробляється специфічний білок інтерферон. Його утворення в організмі пов’язано з комплементарною взаємодією двох неалельних генів, розташованих у різних хромосомах.
Гемоглобін дорослої людини містить чотири поліпептидні ланцюги, кожен з яких кодується окремим незалежним геном. Отже, для синтезу молекули гемоглобіну необхідна наявність чотирьох комплементарних генів.
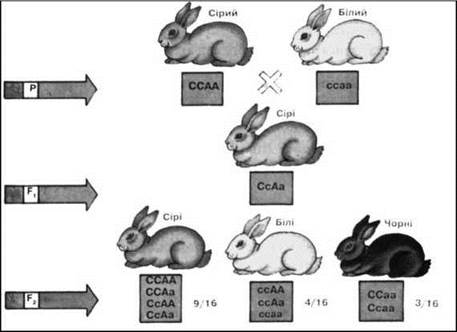
Рис. 20. Комплементарна взаємодія неалельних генів у кролів.
У тварин (мишей, щурів, кролів) для формування темного забарвлення шерсті необхідна присутність двох домінантних неалельних генів (С і А), один з яких визначає наявність пігменту, а інший - його розподіл по волоссю. Якщо один із генів перебуває у гомозиготному рецесивному стані (сс), то пігмент не утворюється і народжуються білі особини (альбіноси).
Епістаз - взаємодія неалельних генів, при якій один ген пригнічує дію іншого, неалельного, гена. Перший ген називається епістатичним, або супресором (інгібітором), інший, неалельний, ген - гіпостатичним. Якщо епістатичний ген - домінантний, епістаз називають домінантним (А>В). І, навпаки, якщо епістатичний ген рецесивний, епістаз - рецесивний (аа>В або аа>вв). Взаємодія генів при епістазі протилежна комплементарності.
У курей домінантний алель С одного гена зумовлює розвиток забарвлення пір'я, але домінантний алель І іншого гена є його супресором. Тому кури з генотипом І-С- — білі, а з генотипами ііСС і ііСс - забарвлені. У схрещуванні білих курей (ІІСС х іісс) гібриди першого покоління F1(IiCc) виявляться білими, але при схрещуванні F1 між собою в другому поколінні F2 відбудеться розщеплення за фенотипом у співвідношенні 13:3. З 16 особин 3 будуть забарвлені (ііСС, ііСс), тому що в них відсутній домінантний ген-супресор і є домінантний ген забарвлення. Інші 13 особин будуть білими. Даний приклад ілюструє явище домінантного епістазу.
Прикладом рецесивного епістазу може бути бомбейський феномен - незвичайне успадкування груп крові системи АВО, вперше виявлене в одній індійській сім'ї. У родині, де батько мав групу крові І (0), а мати - III (В), народилася дівчинка з групою І (0), вона одружилася з чоловіком з групою крові II (А) і в них народилося дві дівчинки: одна з групою крові IV (АВ), інша - з І (0). Народження дівчинки з IV (АВ) групою крові в родині, де батько мав II (А), а мати - І (0) було незвичайним. Генетики пояснили цей феномен так: дівчинка з групою IV (АВ) успадкувала алель Iавід батька, а алель Ів - від матері, але в матері алель Ів фенотипово не виявлявся, тому що в її генотипі був присутній рідкісний рецесивний епістатичний ген s у гомозиготному стані, який пригнічував фенотиповий прояв алеля Ів.
При епістатичній взаємодії генів розщеплення за фенотипом у F2 складає 13:3; 12:3:1 або 9:3:4 та ін.
Гіпостаз - взаємодія неалельних генів, при якій домінантний ген однієї алельної пари пригнічується епістатичним геном з іншої алельної пари. Якщо ген А пригнічує ген В (А>В), то по відношенню до гена В взаємодія неалельних генів називається гіпостазом, а по відношенню до гена А - епістазом.
Полімерія - взаємодія неалельних генів, при якій одну і ту саму ознаку визначають кілька домінантних неалельних генів, які діють на цю ознаку однозначно, в однаковій мірі, підсилюючи її прояв. Такі гени називають полімерними (множинними, полігенами) і позначають однією літерою латинського алфавіту, але з різними цифровими індексами. Наприклад, домінантні полімерні гени - А1, А2, А3 і т.д., рецесивні - а1, а2, а3 і т.д. Відповідно позначають генотипи - А1А2А3; а1а2а3а.,. Ознаки, які контролюються кількома генами (полігенами), називають полігенними, а успадкування цих ознак - полігенним, на відміну від моногенного, де ознака контролюється одним геном. Явище полімерії вперше описав у 1908 р. шведський генетик Г. Нільссон-Еле при вивченні успадкування кольору зерна в пшениці.
Забарвлення насіння пшениці, вівса та ін. може бути від червоного кольору до білого. Інтенсивність забарвлення залежить від числа домінантних генів у генотипі. Вміст всіх чотирьох домінантних алелей (А1А1А2А2) визначає темно-червоне забарвлення, відсутність домінантних генів (а1а1а2а2) - біле, а генотипи з трьома, двома або одним домінантним генами зумовлюють перехідне забарвлення від світло-червоного до рожевого і білого (А1А1А2а2 або А1а1А2А2 - світло-червоного кольору, А1А1а2а2 або а1а1А2А2 - рожевого кольору, а1а1b2b2 - білого кольору). Розщеплення за фенотипом 15:1.
Полімерія буває кумулятивною і некумулятивною. При кумулятивній полімерії кожний ген окремо має слабку дію (слабку дозу), але кількість доз всіх генів у кінцевому результаті сумується, так що ступінь вираження ознаки залежить від числа домінантних алелей. Сумування доз полімерних генів (аддитивність) забезпечує існування неперервних рядів кількісних змін.
За типом полімерії в людини успадковуються ріст, маса тіла, колір шкіри, розумові здібності, величина артеріального тиску, а також у природі успадковується урожайність, яйценосність, молочність та ін. Так, пігментація шкіри в людини визначається 4-6 парами полімерних генів. У генотипі корінних жителів Африки наявні переважно домінантні алелі (Р1Р1Р2Р2Р3Р3Р4Р4), у представників європеоїдної раси - рецесивні (р1р1р2р2р3р3р4р4). Від шлюбу негра і білої жінки народжуються діти з проміжним кольором шкіри - мулати (Р1р1Р2р2Р3р3Р4р4). Якщо подружжя - мулати, то можливе народження дітей з пігментацією шкіри від максимально світлої до максимально темної.
Полігенно в типових випадках успадковуються кількісні ознаки. Проте в природі існують приклади полігенного успадкування якісних ознак, коли кінцевий результат не залежить від числа домінантних алелей у генотипі - ознака або проявляється, або не проявляється (некумулятивна полімерія).
Плейотропія (від грец. πλείων - більш численний і τροπή - напрямок, поворот) - залежність кількох ознак від одного гена, або множинна модифікуюча дія одного гена. Не буває однозначного співвідношення між генотипом і фенотипом. Один і той же ген може діяти на різні ознаки організму. Так, у вищих рослин ген, який визначає забарвлення квіток контролює забарвлення і стебла. У людини домінантний ген, який зумовлює появу арахнодактилії («павучі пальці») викликає одночасно порушення розвитку сполучної тканини, аномалію кришталика ока і вади серця - аневризму аорти (синдром Марфана). Інший приклад, людина може бути носієм гена, який зумовлює відсутність потових залоз. Одночасно цей ген призводить до відсутності деяких зубів та ін. Прояв плейотропії залежить від того, на якій стадії онтогенезу він діє, - чим раніше, тим більший ефект.
Плейотропна дія може бути первинною і вторинною. При первинній плейотропії ген одночасно проявляє свій множинний ефект. Наприклад, при хворобі Хартнепа мутація гена призводить до порушення всмоктування амінокислоти триптофану в кишках і його реабсорбції у ниркових канальцях. При цьому вражаються одночасно мембрани епітеліальних клітин кишок і ниркових канальців з розладами травної і видільної систем. При вторинній плейотропії є один первинний фенотипний прояв гена, услід за яким розвивається ступінчастий процес вторинних змін, які призводять до множинних ефектів. Так при серпоподібно клітинній анемії у гомозигот спостерігається кілька патологічних ознак: анемія, збільшена селезінка, ураження шкіри, серця, нирок, мозку. Тому гомозиготи за геном серпоподібноклітинної анемії гинуть, як правило, ще в дитячому віці. Всі ці фенотипові прояви складають ієрархію вторинних проявів. Першопричиною, безпосереднім фенотипним проявом дефектного гену є аномальний гемоглобін і еритроцити серпоподібної форми. Внаслідок цього відбуваються послідовно інші патологічні процеси: злипання і руйнування еритроцитів, анемія, дефекти у серці, нирках, мозку. Ці патологічні ознаки є вторинними. Більш розповсюджена вторинна плейотропія.
Множинний алелізм
У дослідах Г. Менделя гени існували лише в двох формах - домінантній і рецесивній. Але більшість генів представлена не двома, а більшим числом алелей. Крім основних алелей (домінантного і рецесивного) існують ще проміжні алелі. Серію алелей (три і більше) одного гена називають множинними алелями, а саме явище - множинним алелізмом. Множинні алелі виникають внаслідок багаторазових мутацій одного і того самого локусу хромосоми. У генотипі диплоїдного організму представлені лише два алелі одного гена, у популяції число їх практично не обмежене. Особливість взаємодій між множинними алелями та, що їх можна розмістити в один послідовний ряд, в якому кожний член буде домінантним по відношенню до всіх наступних і рецесивним по відношенню до попередніх. Наприклад, у кроликів суцільне темне забарвлення зумовлене домінантним алелем А, гомозиготні рецесивні тварини (аа) - білі. Але існує ще кілька алелей цього гена, які мають свій власний фенотип у гомозиготному стані - шиншиловий (achach) і гімалайський (ahah). Шиншилові кролики - сірі, гімалайські - білі, але кінчики вух, хвоста, ніг і носа забарвлені. Вся серія алелей може бути записана у вигляді послідовного ряду A>ach>ah>a. Темний колір будуть мати кролики з генотипами АА, Aach, Aah, Аа, шиншиловий - з генотипами achach, achah, acha, гімалайський - з генотипами ahah, aha. Значення: множинний алелізм збільшує генофонд популяції, її генотиповий і фенотиповий поліморфізм, що має значення для еволюції.
Успадкування груп крові за системою АВO у людини пов’язано з серією множинних алелей.
Успадкування груп крові системи АВО та MN; генетика груп крові
Відкриття АВО-системи груп крові належить К. Ландштейнеру (1901). Система груп крові АВО у людини успадковується за типом множинних алелей одного автосомного гена, розташованого у хромосомі 9, локус якого позначається літерою І (від слова ізогемаглютиноген). Вивченням характеру успадкування різних груп крові АВО-системи встановлено, що вони визначаються різним поєднанням трьох алелей однієї алеломорфної групи генів, які позначають IA, Ib та I0. Вони визначають чотири фенотипи: з першою групою І (0), другою II (А), третьою III (В), четвертою IV(АВ) (табл.).
Групи крові системи АВО
Групи крові (фенотипи) |
Генотипи |
Антигени еритроцитів |
Антитіла плазми крові (аглютиніни) |
І (0) |
І0І0 |
немає |
α і β |
II (А) |
ІАІА, ІАІ0 |
А |
β |
III (В) |
ІВІВ, ІВІ0 |
В |
αа |
IV (АВ) |
ІАІВ |
А, В |
немає |
Кожен з фенотипів відрізняється специфічними білками-антигенами, які містяться у еритроцитах, і антитілами - зосереджуються у сироватці крові. Фенотип І (0) зумовлений відсутністю у еритроцитах антигенів А і В і наявністю у сироватці крові антитіл α і β. Фенотип ІІ (А) характеризується наявністю у еритроцитах антигена А і антитіл β у сироватці крові. Фенотип ІІІ (В) пов’язаний з наявністю у еритроцитах антигена В і антитіл α у сироватці крові. Фенотип ГУ (АВ) залежить від наявністі у еритроцитах антигенів А і В та відсутності у сироватці крові антитіл α і β.
Разом антиген А і антитіло а не містяться ніколи, як і антиген В з антитілом р. При взаємодії антигенів з однойменними антитілами відбувається склеювання еритроцитів (аглютинація), що свідчить про несумісність крові донора і реципієнта. При переливанні крові необхідно, щоб антигени донора не зустрілися з однойменними антитілами реципієнта. Оскільки перша група не має антигенів, то люди з такою кров'ю називаються універсальними донорами, а люди з четвертою групою - універсальними реципієнтами.
Успадкування двох алелей з трьох можливих підкоряється менделівським закономірностям. Групи крові II (А) і III (В) успадковуються за автосомно-домінантним типом, І (0) група-за автосомно-рецесивним. Г ени Iа і IB по відношенню до гена I0 ведуть себе домінантно. Якщо батьки мають групу крові II (А), то їхні діти можуть мати II (А) і І (0), але не III (В) і не IV (АВ). Четверта група крові (АВ) успадковується не за правилами Г. Менделя, а за типом кодомінування. Оскільки групи крові генетично зумовлені і не змінюються протягом життя, то їх визначення може допомогти у випадку спірного батьківства. При цьому необхідно пам'ятати, що за групою крові не можна встановити, що саме цей мужчина є батьком дитини. Можна лише сказати, що він міг бути батьком дитини чи батьківство виключене.
В осіб з IV (АВ) групою крові в 0,1-0,2 % випадків спостерігається особливе положення генів - цис-положення, коли обидва гени Iаі Ів знаходяться в одній хромосомі. Тоді в шлюбі такої людини з особою, що має І (0) групу крові, можливе народження дітей з І (0) групою крові, що необхідно враховувати при медико-генетичному консультуванні, проведенні судово-медичної експертизи.
Люди з генотипом IaIa фенотипово зовсім не різняться від людей з генотипом IaI0, але мають особливості їх діти. У дітей від шлюбу, в якому один із батьків має генотип IaI0, а інший І0І0, половина дітей має фенотип А (при генотипі IaI0), і половина - фенотип 0. Якщо один з батьків має генотип IaIa, а другий I0I0, то всі діти мають фенотип А (при генотипі IaI0). Така ж різниця спостерігається й у людей з генотипами ІВІВ і ІВІ0.
Нащадки, очікувані від шлюбу, відповідно до груп крові батьків
Типи поєднання |
Групи крові батьків |
Групи крові в дітей |
1 |
00x00 |
00 |
2 |
00хА0 |
А0, 00 |
00хАА |
А0 |
|
3 |
00хВ0 |
В0, 00 |
00хВВ |
В0 |
|
4 |
А0хА0 |
АА, А0, 00 |
ААхА0 |
АА, А0 |
|
ААхАА |
АА |
|
5 |
А0хВ0 |
АВ, А0, В0, 00 |
ААхВ0 |
АВ, А0 |
|
А0хВВ |
АВ, В0 |
|
ААхВВ |
АВ |
|
6 |
В0хВ0 |
ВВ, В0, 00 |
В0ХВВ |
ВВ, В0 |
|
ВВхВВ |
ВВ |
|
7 |
00хАВ |
А0, В0 |
8 |
А0хАВ |
АА, АВ, А0, В0 |
ААхАВ |
АА, АВ |
|
9 |
В0хАВ |
АВ, ВВ, А0, В0 |
ВВхАВ |
АВ, ВВ |
|
10 |
АВхАВ |
АА, АВ, ВВ |
Примітка. У межах кожного з десяти типів поєднань генотип групи крові можна розрізнити тільки на підставі вивчення нащадків.
Фенотипові прояви АВ0-системи груп крові відносяться до найбільш стійких ознак і за життя людини ніколи не зазнають змін.
Виділяють понад 20 різних систем крові за наявністю антигенів, у тому числі за резус-фактором та системою MN.
При успадкуванні групи крові за системою MN, відкритою у 1927 році, також має місце кодомінування. Ця система визначається двома алелями: ІМ і IN. Обидва алелі кодомінантні, тому існують люди з генотипом ІМІМ (у фенотипі вони мають фактор М), ININ (у фенотипі вони мають фактор N), IMIN (у фенотипі у них обидва фактори М і N). У сироватці крові людей з тим чи іншим фенотипом за цією системою груп крові немає антитіл до відповідних антигенів, як це має місце у системі АВ0. Тому при переливанні крові ця система може не враховуватися. Серед європейців генотип ІМIМ зустрічається приблизно у 36 %, ININ - у 16 %, IMIN - у 48%.
Успадкування резус-фактора
Резус-фактор - білок (антиген), названий так тому, що вперше (1940 р.) був виділений з еритроцитів мавпи макака-резус (Macacus resus), а потім у людини. Близько 85 % європейців здатні його синтезувати і становлять резус-позитивну групу (Rh), 15 % - нездатні і називаються резус-негативними (Rh-). Резус-фактор зумовлений трьома домінантними тісно зчепленими генами (С, D, Е), розміщеними в першій хромосомі. Успадковуються вони як при моногібридному схрещуванні. Основна роль належить антигену D, якщо він визначається, то кров належить до резус-позитивної (DD або Dd), якщо не визначається - то до резус- негативної (dd). Резус-фактор необхідно враховувати при переливанні крові і трансплантації, тому що на нього в організмі виробляються антитіла. Резус-фактор може бути причиною резус-конфлікту між матір'ю і плодом. При шлюбі жінки, що має резус-негативну кров, з чоловіком, який є резус-позитивною гомозиготою, усі діти будуть резус-позитивними, а при його гетерозиготності - 50 % резус-позитивні і 50 % резус-негативні.
Конфлікт виникає в тому випадку, коли мати має резус-негативну кров, а дитина одержала домінантний алель D від батька і є резус-позитивною. Кров матері і плода не змішується. Тому перша вагітність завершується нормально. Але під час народження першої дитини, коли плацента відшаровується, еритроцити дитини попадають в організм матері, де на резус-антиген утворюються антитіла. При наступній вагітності ці антитіла через бар'єр плаценти проникають у кров плода, з'єднуються з резус-антигеном, викликаючи склеювання еритроцитів та їх лізис (еритроблазтоз, або гемолітична хвороба новонароджених). Причому з кожними наступними пологами захворювання в дітей виявляється в більш тяжкій формі. Якщо резус-негативній дівчині до шлюбу було зроблено переливання резус- позитивної крові, то вже перша дитина (якщо вона резус-позитивна) буде нежиттєздатною. Тому навіть одноразове переливання резус-позитивної крові дівчатам з резус-негативною кров'ю абсолютно не припустиме.
В Японії, Кореї, Китаї, Індії гемолітична хвороба новонароджених зустрічається дуже рідко. Пояснюється це тим, що серед них дуже низька частота Rh(-) (від 0 до 1,5 %). Рідко зустрічається Rh(-) група крові у ескімосів, евенків. У австралійських аборигенів резус-конфлікт у вагітних не існує, так як частота гена Rh(+) у них складає 100 %.
Гемолітична хвороба новонароджених описана понад 400 років назад. Вона виникає при несумісності не тільки за резус-системою, але й за системою АВ0: найбільш часто, коли в матері І (0) група, а в дитини II (А) або III (В).
Імуногенетика
Імуногенетика - наука, яка вивчає закономірності успадкування антигенної специфічності і генетичну зумовленість імунних реакцій. В організмі людини є головний комплекс гістосумісності (Major Histocompatibility Complex - МНС) - система антигенів, яка властива певній людині і зумовлює тканинну несумісність донора і реципієнта. Антигени МНС добре виражені на поверхні лейкоцитів периферичної крові, тому головний комплекс гістосумісності в людини одержав назву HLA-система (Human Leucocyte Antigen - людські лейкоцитарні антигени). Синтез цих антигенів контролюється групою тісно зчеплених генів, які розміщені в 6-й парі хромосом. Тут є локуси А, В, С, D1, D2 R. Кожний ген має по декілька алелей. Відомо понад 20 алелей гена А, близько 50 алелей - гена В, 8 - гена С. Комбінація цих алелей створює різноманітність генотипів людей за антигенами в популяціях. Для визначення у людини HLA-антигенів використовуються панелі сироваток у мікротестах токсичності. Визначення антигенів гістосумісності має велике значення для пересадки органів і тканин, створення банків даних про антигени реципієнтів і донорських органів. Крім того, є дані про зв'язок між деякими захворюваннями і певними HLA-антигенами. Наприклад, антигени HLA-B27 зустрічаються в 70-100 % людей, хворих на анкілозируючий спондиліт, і в 3-12 % здорових. Гени головного комплексу гістосумісності контролюють функціонування імунної системи, забезпечують утворення трансплантаційного імунітету.