ОСНОВИ МЕДИЧНОЇ БІОЛОГІЇ - 2012
Зчеплене успадкування. Генетика статі
Незалежне комбінування генів може здійснюватись лише за умов, що гени знаходяться в різних парах хромосом. У кожного організму кількість генів, які незалежно комбінуються при мейозі, обмежена кількістю пар хромосом. Але кількість ознак організму, які контролюються генами, дуже велика, а кількість пар хромосом невелика. Якщо допустити, що в кожній хромосомі не один ген, а більше, тоді яким чином успадковуються гени, локалізовані в одній хромосомі? Відповідь на ці питання одержав Т. Морган в дослідах на плодовій мушці дрозофілі. Гени — складові частини хромосом. Так як генів більше, ніж хромосом, можна передбачити, що в одній хромосомі буде не один ген, а кілька, які успадковуються разом. Таке явище було назване Т. Морганом зчепленням генів. Успадкування генів, які знаходяться в одній хромосомі, що обмежує вільне їх комбінування, називається зчепленням генів.
Гени, розташовані в одній хромосомі, називаються зчепленими і успадковуються переважно разом (зчеплене успадкування). Усі гени однієї хромосоми утворюють одну групу зчеплення. Гомологічні хромосоми мають однакові групи зчеплення, тому в кожному організмі буде стільки груп зчеплення, скільки пар гомологічних хромосом.
Ознаки, гени яких належать до однієї групи зчеплення, не підкоряються принципу незалежного розподілу при дигібридному схрещуванні (АаВb х АаВb) у співвідношенні 9:3:3:1 і при дигібридному аналізуючому схрещуванні (АаВb х aabb) у співвідношенні 1:1:1:1. Разом зчеплені гени передаються не завжди.
Явище зчеплення ознак відкрили в 1906 р. В. Бетсон і Р. Пеннет у дослідах із запашним горошком при схрещуванні двох його рас, які відрізнялися двома парами ознак - формою пилку та кольором квітів. Замість очікуваного розщеплення в другому поколінні F2 (9:3:3:1) спостерігалося розщеплення в співвідношенні, близькому до 3:1. Ознаки, таким чином, не проявляли незалежного успадкування.
Явище зчеплення генів проаналізував Т. Морган. У дрозофіли алель сірого кольору тіла (В) домінує над алелем чорного кольору тіла (b), алель нормальної довжини крил (V) - над алелем зачаткових крил (v). Схрещування гомозиготної сірої мухи з нормальними крилами (BBVV) і чорної мухи із зачатковими крилами (bbvv) дало в першому поколінні F1 гібридів з сірим тілом і нормальними крилами (BbVv), що підтверджувало закон одноманітності гібридів першого покоління, встановлений Г. Менделем. Результати досліду не залежали від статі рецесивної гомозиготи. Далі було проведено два аналізуючих дигібридних схрещування. У першому з них самець був дигібридом F, (сіре тіло і нормальні крила), а самка - гомозиготою за рецесивними алелями (чорне тіло і зачаткові крила). Від цього схрещування було одержано два фенотипових класи, аналогічні вихідним батьківським формам, у рівному співвідношенні: 50% сірих з нормальними крилами (BbVv) і 50% чорних із зачатковими крилами (bbvv). У другому дигібридному аналізуючому схрещуванні самка була дигібридом F1 (сіра з нормальними крилами), а самець - рецесивною гомозиготою (чорний із зачатковими крилами). Від цього схрещування було одержано чотири фенотипових класи в співвідношенні:
1) сірі з довгими крилами (BbVv) 41,5 %;
2) сірі із зачатковими крилами (Bbvv) 8,5 %;
3) чорні з довгими крилами (bbVv) 8,5 %;
4) чорні із зачатковими крилами (bbvv) 41,5 % (рис.)
Результати обидвох аналізуючих дигібридних схрещувань не співпадали з очікуваним співвідношенням фенотипів - 25 % сірих з нормальними крилами, 25 % сірих із зачатковими крилами, 25 % чорних з нормальними крилами і 25 % чорних із зачатковими крилами, як це має місце при незалежному успадкуванні (Г. Мендель). Відхилення від очікуваного розщеплення (1:1:1:1) Морган пояснив тим, що гени досліджуваних ознак (В і V) розташовані в одній хромосомі і успадковуються разом (зчеплено). Сила зчеплення між генами обернено пропорційна відстані між ними в хромосомі (правило або закон Моргана).
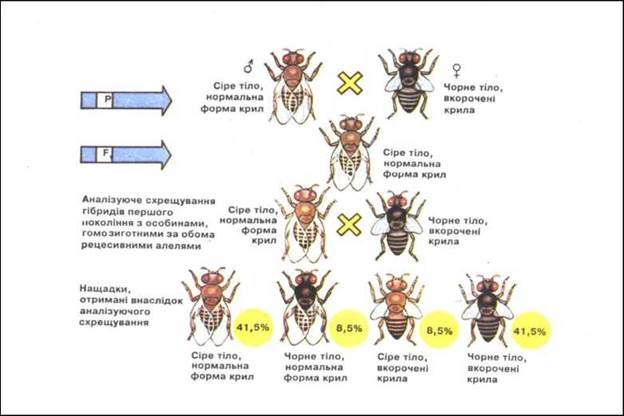
Рис. 21. Зчеплене успадкування в дрозофіли.
Зчеплення генів може бути повним і неповним. У першому аналізуючому схрещуванні ![]() мало місце повне зчеплення генів. У другому аналізуючому схрещуванні
мало місце повне зчеплення генів. У другому аналізуючому схрещуванні ![]() число особин з фенотипами, які повторюють фенотипи батьків, переважало (83 %) і теж вказувало на тісне зчеплення алелей В і V. Появу в меншій кількості (17%) потомства з фенотипами, в яких поєднувалися ознаки обидвох батьків, Т. Морган, опираючись на відкриття хіазм у мейозі Ф. Янсеном (1909), пояснив порушенням зчеплення в результаті перехресту хромосом у точці між генами В і V. Процес перехресту Морган назвав кросинговером. Відмінності в результатах 1-го аналізуючого схрещування, коли дигібридом був самець, і 2-го аналізуючого схрещування, коли дигібридом була самка, пояснюються особливістю біології дрозофіли, яка полягає у відсутності кросинговеру в гаметогенезі самців. Тому дигібридний самець (BbVv) утворює тільки некросоверні гамет (BV і bv) двох сортів по 50 % кожного сорту, а дигібридна самка (BbVv) - 83 % некросоверних гамет двох сортів (41,5 % BV і 41,5 % bv) і 17 % кросоверних гамет двох сортів (8,5 % Bv і 8,5 % bV). Кросоверними називаються гамети, в процесі утворення яких відбувся кросинговер, некросоверні гамети утворилися без кросинговеру. Відповідно особини, які виникли з участю кросоверних гамет, називаються кросоверами, а утворені без них - некросоверами. Отже, в досліді Т. Моргана кросоверів у потомстві було 17%, некросоверів - 83%.
число особин з фенотипами, які повторюють фенотипи батьків, переважало (83 %) і теж вказувало на тісне зчеплення алелей В і V. Появу в меншій кількості (17%) потомства з фенотипами, в яких поєднувалися ознаки обидвох батьків, Т. Морган, опираючись на відкриття хіазм у мейозі Ф. Янсеном (1909), пояснив порушенням зчеплення в результаті перехресту хромосом у точці між генами В і V. Процес перехресту Морган назвав кросинговером. Відмінності в результатах 1-го аналізуючого схрещування, коли дигібридом був самець, і 2-го аналізуючого схрещування, коли дигібридом була самка, пояснюються особливістю біології дрозофіли, яка полягає у відсутності кросинговеру в гаметогенезі самців. Тому дигібридний самець (BbVv) утворює тільки некросоверні гамет (BV і bv) двох сортів по 50 % кожного сорту, а дигібридна самка (BbVv) - 83 % некросоверних гамет двох сортів (41,5 % BV і 41,5 % bv) і 17 % кросоверних гамет двох сортів (8,5 % Bv і 8,5 % bV). Кросоверними називаються гамети, в процесі утворення яких відбувся кросинговер, некросоверні гамети утворилися без кросинговеру. Відповідно особини, які виникли з участю кросоверних гамет, називаються кросоверами, а утворені без них - некросоверами. Отже, в досліді Т. Моргана кросоверів у потомстві було 17%, некросоверів - 83%.
Кросинговер (перехрест хромосом) - взаємний обмін ділянками гомологічних хромосом. Закономірно відбувається в профазі першого мейотичного поділу на стадії пахінеми при гаметогенезі (сперматогенезі та овогенезі). Гомологічні хромосоми (материнська і батьківська) профази І найтісніше прилягають одна до одної (кон'югують). Кожна з них представлена двома хроматидами. Перед тим як розійтися, хроматиди утворюють Х-подібні фігури (хіазми), у точках перехресту хроматиди відриваються, а потім з'єднуються, але вже материнська не з материнською, а материнська з батьківською. Обмін генетичним матеріалом відбувається, як правило, між несестринськими хроматидами гомологічних хромосом, але може відбуватися і між сестринськими. Кросинговер порушує зчеплення генів. У результаті виникають рекомбінантні хромосоми з новими поєднаннями генів, нові групи зчеплення, що веде до появи потомства з новими комбінаціями батьківських генів. Кросинговер має значення для еволюції як один з механізмів виникнення комбінативної мінливості.
Крім мейотичного кросинговеру, який закономірно відбувається в мейозі при утворенні гамет, іноді спостерігається обмін генетичним матеріалом між хроматидами гомологічних хромосом у соматичних клітинах (мітотичний кросинговер). Так, автосомно-рецесивна мутація людини, відома як синдром Блюма, супроводжується цитологічною картиною, яка нагадує кон'югацію гомологічних хромосом і навіть утворення хіазм.
Частоту кросинговеру вимірюють відношенням числа кросоверів до загального числа особин у потомстві від аналізуючого схрещування і виражають у відсотках. Частота кросинговеру між генами прямо пропорційна відстані між ними. Чим ближче між собою розміщені гени, тим менше між ними можливих точок кросинговеру і, навпаки, чим далі між собою розміщені гени, тим більше між ними можливих точок кросинговеру. Кросинговер між двома генами може відбуватися не тільки в одній, але і в двох (подвійний кросинговер) і навіть більшому числі точок.
Частота кросинговеру відбиває ступінь зчеплення генів і для даних двох генів, за однакових умов, завжди постійна. Для інших генів цієї ж хромосоми частота кросинговеру обов'язково інша, але теж постійна: від часток відсотку до майже 50 %. Постійність відсотку кросинговеру між генами використовується як показник відносної відстані між ними. За одиницю відстані між генами прийнята одна морганіда. Вона відповідає відстані, на якій частота кросинговеру дорівнює 1 %. У досліді Моргана кросоверів у потомстві було 17 %. Звідси, частота кросинговеру між генами В і V дорівнює 17 %, відстань між генами - 17 морганід. На основі цих даних Т. Морган та його співробітники розробили положення про лінійне розміщення генів у хромосомах і запропонували принцип побудови хромосомних карт.
Основні положення хромосомної теорії спадковості
Хромосомну теорію спадковості сформулювали і експериментально обгрунтували в дослідах на плодовій мушці дрозофілі (Drosophila melanogaster), проведених у 1910-1925 pp. американський генетик Т. Морган та його школа - А. Стертевант, Г. Меллер, К. Бріджес. Основні положення хромосомної теорії спадковості:
1. Гени розміщаються в хромосомах по довжині в лінійному порядку; різні хромосоми містять неоднакове число генів; набір генів кожної з негомологічних хромосом - унікальний.
2. Алельні гени займають певні й ідентичні локуси (місця) гомологічних хромосом.
3. Гени, розташовані в одній хромосомі, утворюють групу зчеплення, завдяки чому має місце зчеплення деяких ознак, які разом (зчеплено) передаються нащадкам. Кількість груп зчеплення дорівнює гаплоїдному набору хромосом. Зчеплення не абсолютне.
4. Під час мейозу, який відбувається тільки при утворенні гамет, диплоїдне число хромосом зменшується вдвічі (гаплоїдне число). Це відповідає закону розщеплення, за яким генетичний матеріал обох батьків має розщепитися і потрапити в різні гамети.
5. Згідно із законом незалежного розподілу батьківські й материнські набори незчеплених генів розщеплюються незалежно один від одного. Якщо незчеплені гени розташовані в різних хромосомах, то під час мейозу материнські й батьківські хромосоми повинні розподілитися між гаметами випадково.
6. Між генами гомологічних батьківських і материнських груп зчеплення можуть відбуватися, завдяки кросинговеру, реципрокні рекомбінації. Цьому відповідає утворення хіазм під час кон’югації гомологічних хромосом у мейозі (генетичний кросинговер).
7. Сила зчеплення між генами обернено пропорційна відстані між ними. Чим ближче розташовані гени в одній хромосомі, тим сильніше їх зчеплення, тим у меншій мірі виникатимуть рекомбінації між ними, і навпаки. Відстань між генами вимірюється у відсотках кросинговеру. Один відсоток кросинговеру відповідає одній морганіді.
8. Кожний біологічний вид характеризується специфічним набором хромосом - каріотипом. Дослідження хромосомного механізму визначення та успадкування ознак, зчеплених зі
статтю, зчеплення генів, генетичних карт з цитологічними картами хромосом - все це покладено в основу хромосомної теорії спадковості, на яку спирається весь розвиток генетики.
Генетичні карти хромосом
Ґрунтуючись на знанні теорії лінійного розташування генів у хромосомі та на підставі кросинговеру, визначивши відстань між ними, вдалося встановити їх послідовність у хромосомі. А.Стертевант склав першу карту розподілу генів в одній із хромосом дрозофіли. Потім були складені такі карти і для інших хромосом. За одиницю відстані між генами приймається 1% кросинговеру, або 1 морганіда.
Генетична карта - це відрізок прямої, на якій позначено порядок розташування генів і вказано відстань між ними у відсотках кросинговеру. Вона будується на основі результатів аналізуючого схрещування.
Картування проводиться з метою, щоб дізнатися, в якій парі хромосом і на якій генетичній відстані (коефіцієнт рекомбінації), або в якій частині хромосоми, розташовані гени.
Побудовані генетичні карти для багатьох об’єктів. Серед покритонасінних рослин краще всього вивчено генетичні карти кукурудзи, ячменю, рису, томатів та ін. У тварин такі карти складено для дрозофіли, а серед ссавців - для миші. Доведено, що такий розподіл генів у хромосомах є загальнобіологічною закономірністю.
У людини аналіз зчеплення генів класичними методами, розробленими на дрозофілі, неможливий, внаслідок неможливості експериментальних шлюбів. В останні роки розроблено комплекс методів картування генів на хромосомах людини. Так, для вивчення груп зчеплення і складання карт хромосом використовують понад 15 методів.
Картування хромосом у людини проводиться шляхом аналізу родоводів та генетичного аналізу гібридних соматичних клітин.
Тривалий час були відомі тільки три автосомні групи зчеплення і Х-хромосомна. Новий період розпочався з 1968 р., коли вдалося встановити локалізацію гена групи крові Даффі у хромосомі 1. У людини відомі всі 24 групи зчеплення.
Геном людини містить від 35000 до 40000 різних генів, 16000 з яких тепер картовані на скелетній карті генома, складеній за допомогою 1000 високополімерних ДНК-маркерів (J. Schuler et al., 1996).
Виявлення груп зчеплення у хромосомах дає можливість побудови хромосомних карт, коли на хромосомі розміщення генів позначають у вигляді цяточок. Найбільш повно побудована карта Х-хромосоми. Локуси, зчеплені з Х-хромосомою (а їх понад 200), віднесені до цієї хромосоми спираючись на аналіз родоводів (багато з них підтверджені методами гібридизації соматичних клітин).
Методи побудови карт хромосом ґрунтуються на передбаченні ймовірності кросинговеру по всій довжині хромосоми і випадкових обмінах між різними хроматидами.
Для картування хромосом застосовують гібридизацію соматичних клітин людини з клітинами різних тварин, переважно з клітинами миші. При культивуванні таких гібридних клітин на живильних середовищах відбувається постійна втрата хромосом людини. Коли в гібридній клітині залишається тільки одна хромосома від людського каріотипу, її можна ідентифікувати шляхом диференціального забарвлення. Визначивши потім вміст певного ферменту в даній клітині, можна стверджувати, що локус для цього ферменту розташований у цій хромосомі.
Незважаючи на певні труднощі при побудові генетичних карт і на їх відносність, детальна розробка карт - один з провідних методів аналізу поведінки і тонкої будови хромосом.
Уже відомі 24 групи зчеплення у людини (22 пари автосом, Х, Y). Картована значна кількість генів на хромосомах людини.
Складання точних карт хромосом є необхідною передумовою подальшого прогресу теоретичної генетики людини. Це має виняткове значення для виявлення, диференціальної діагностики спадкових хвороб плода, для ранньої діагностики цих хвороб, встановлення носіїв генетичних зрушень, які перебувають у безсимптомній фазі, для визначення медико-генетичного прогнозу.
Сучасний стан дослідження геному людини
Міжнародна програма «Геном людини» була сформульована на початку 90-х років ХХ століття. Внаслідок копіткої роботи до початку 2003р. геном людини повністю секвенований, тобто повністю прочитана послідовність трьох мільярдів пар основ, з яких побудована ДНК всіх 23 пар хромосом людини. Генетична довжина геному людини складає 3000 сМ (сантиморганіда, генетична відстань, яка дорівнює 1% кросинговеру). Розв’язання програми «Геном людини» сприяло створенню генетичної карти, відтворенню цитогенетичної карти геному.
Генетична карта передбачає встановлення послідовності розміщення генетичних маркерів з відстанню не більше 1 см вздовж усіх хромосом. Така генетична карта дозволяє картувати будь- який ген, встановити відносну відстань між локусами.
Винятковим успіхом програми «Геном людини» було створення інтегральних (фізичних) карт геному. Картовано близько 40000 кодуючих послідовностей. Загальне число генів, очевидно,складатиме 30500-40000.
На сьогодні весь геном людини клонований у вигляді великих фрагментів, які перекривають один одного. Розташування кожного з цих фрагментів на хромосомі визначено з високою точністю. Термін клонування означає, що ген картований, вивчена його структура, знайдена мутація, яка викликає те чи інше захворювання.
Розкриття геному людини сприятиме розвитку нових напрямків медицини, вивченню природи спадкових і злоякісних хвороб, розробці генної і клітинної терапії.
Нехромосомна (цитоплазматична) спадковість
Теорія спадковості Т. Моргана називається хромосомною. Цим підкреслюється провідна роль хромосом ядра в явищах спадковості. Проте спадковість пов'язана також і з цитоплазмою. На відміну від хромосомної (ядерної) ця форма спадковості називається нехромосомною, або цитоплазматичною. Вона забезпечується генами, які локалізовані поза ядром клітини - у мітохондріях і пластидах, які мають власну ДНК і здатні до автономної (незалежно від ядра) редуплікації. Сукупність генів цитоплазми називається плазмоном, а самі гени - плазмагенами. Плазмагени за своїми властивостями схожі з ядерними генами. Вони здатні до редуплікацій і мутацій. Цитоплазматична спадковість передається в однобічному напрямі виключно по материнській лінії з цитоплазмою яйцеклітин. За материнським типом успадковуються стійкість до стрептоміцину в хламідомонад, напрямок завитки черепашки прісноводних молюсків (Lymnea), ряболистість у декоративних рослин нічної красуні (Mirabilis jalapa), левових ротиків (Antirrhinum majus), цитоплазматична чоловіча стерильність у кукурудзи. У бактерій цитоплазматична спадковість пов'язана з плазмідами - позахромосомними невеликими молекулами ДНК кільцевої форми. Відомі різноманітні за своєю функцією плазміди. F-плазміди забезпечують здатність бактерій до кон'югації, R-плазміди - множинну резистентність до лікарських засобів, Col-плазміди - синтез коліцинів (білків з летальною активністю).
Основні форми цитоплазматичної спадковості - пластидна і мітохондріальна. Приклад пластидної спадковості - успадкування ряболистості в нічної красуні, інших рослин.
Мітохондріальна спадковість. Кожна мітохондрія має власну ДНК. У порівнянні з ядерними ДНК, мітохондріальні ДНК (mДHК) мають кільцеву форму, відносно невеликі розміри і відповідно невеликий набір генів, не містять інтронів. У людини тДНК містить 16569 пар нуклеотидів. Генетичний код мітохондріальних ДНК відрізняється від універсального: окремі триплети кодують інші амінокислоти, неоднакові нуклеотидні послідовності формують стоп-кодони, частіше в третій позиції зустрічається аденін або цитозин. Більшість мітохондріальних білків кодується ядерними ДНК і синтезується в цитоплазмі, а потім надходить у мітохондрії. І лише приблизно 2 % мітохондріальних білків кодується плазмагенами і синтезується на мітохондріальних рибосомах. З мутаціями мітохондріальних генів пов'язані мітохондріальні хвороби.
Генетика статі
Кожний вид організмів має певне співвідношення статей при народженні у більшості організмів співвідношення між статями в популяції дорівнює 1:1 (або 50% на 50%):
Тому, Г. Мендель та Донкастер пропонували таку гіпотезу генетичного механізму визначення статі — одна стать гетерозиготна (Аа), а інша — гомозиготна за рецесивними алелями і тоді (див. схему) із покоління в покоління буде зберігатися співвідношення між статями 1:1.
Стать успадковується як менделююча ознака. Розщеплення за статтю відбувається як при аналізуючому моногібридному схрещуванні в співвідношенні 1 : 1.
Стать тварин і рослин генетично визначається однією парою хромосом, які отримали назву статевих. Існують чотири основних типи регуляції статі статевими хромосомами.
1. XY-тип, за якого жіноча стать має дві ХХ статеві хромосоми і є гомозиготною (утворює гамети одного типу - Х), а чоловіча стать має чоловічу статеву хромосому - Y, яка відрізняється від Х-хромосоми величиною і формою. Чоловіча стать гетерогаметна (утворює гамети двох типів - Х і Y). Цей тип властивий ссавцям, комахам і більшості дводольних покритонасінних рослин.
2. Х0-тип - жіноча стать має дві Х-хромосоми, а чоловіча тільки одну Х-хромосому.
Х0-тип трапляється серед комах і ссавців.
3. ZW-тип. Жіноча стать має одну жіночу статеву хромосому W і другу, відмінну від неї за формою і величиною, статеву хромосому Z. Тут жіноча стать є гетерогаметною, а чоловіча стать має дві однакових статевих хромосоми Z і гомогаметна. ZW-тип притаманний деяким рибам, метеликам, птахам і дуже рідко трапляється серед рослин.
4. ZO-тип - жіноча стать має тільки одну Z-хромосому і гетерогаметна, а чоловіча - дві Z- хромосоми і гомогаметна. Цей тип відомий тільки в одного з видів ящірки.
Способи визначення статі:
- прогамний тип - стать визначається ще до запліднення (у дафній за оптимальних умов у водоймі розвиваються самки, а при низькій температурі - самці);
- сингамний тип - стать визначається в момент запліднення (у людини при поєднанні двох Х-хромосом формується жіночий організм, а X- та Y-хромосоми - чоловічий);
- епігамний тип - стать визначається після запліднення і формується під впливом умов довкілля (типовим прикладом є морський черв’як Bonellia viridis, самка якого відкладає запліднені яйця у воду, де з них розвиваються личинки. Якщо личинка попадає на хоботок самки, то з неї під дією певних ферментів розвивається великих розмірів самка. Якщо ж личинка опускається на дно водойми або прикріплюється до якогось іншого субстрату - то з неї розвивається дрібних розмірів самець);
Стать визначається також залежно від факту запліднення (у бджіл із заплідненої яйцеклітини формуються самки, а з незаплідненої - самці).
Стать, як і будь-яка інша ознака, генетично детермінована. У людини стать визначається в момент запліднення (сингамно) і залежить від комбінації статевих хромосом від обох батьків у зиготі. Розрізняють гомогаметну і гетерогаметну стать. Гомогаметна - стать, в якій обидві статеві хромосоми однакові та утворює один тип гамет. Гетерогаметна - стать з різними статевими хромосомами та утворює два типи гамет. У людини, ссавців, дрозофіли гомогаметною статтю є жіноча стать, гетерогаметною - чоловіча. Жінка (XX) в мейозі утворює один тип яйцеклітин (X), чоловік (XY) - два типи сперматозоїдів у рівному співвідношенні: половина сперматозоїдів містить X-хромосому, інша половина - Y- хромосому.
Стать майбутнього організму залежить від гетерогаметної батьківської особини, яка утворює два типи гамет: та з цих гамет, яка візьме участь у заплідненні, та і визначить стать нащадка. Народиться дівчинка чи хлопчик, залежить від типу сперматозоїда, бо яйцеклітини за статевою хромосомою однакові. Можливі дві комбінації: 1) якщо яйцеклітину X запліднює Х-сперматозоїд, у зиготі поєднуються дві Х-хромосоми: з такої зиготи розвивається організм жіночої статі XX; 2) якщо яйцеклітину X запліднює Y-сперматозоїд, у зиготі поєднуються X- і Y-хромосоми: з такої зиготи розвивається організм чоловічої статі XY.
Ознаки, гени яких розташовані не в автосомах, а в статевих хромосомах (X і Y), називають зчепленими зі статтю, а успадкування таких ознак - успадкуванням, зчепленим зі статтю. Його вперше встановив Т. Морган при вивченні успадкування кольору очей (червоного і білого) у дрозофіли.
Зверху ліворуч - гени, розташовані в непарній з Y-хромосомою ділянці Х-хромосоми; зверху праворуч - гени, розташовані в непарній ділянці Y-хромосоми; внизу праворуч і ліворуч - гени, розташовані в гомологічних ділянках X- і Y-хромосом.
Першу групу складають гени, повністю зчеплені зі статтю. Вони локалізовані в негомологічній ділянці Х-хромосоми, яка не має гомолога в Y-хромосомі, і передаються виключно через X-хромосому. До числа таких генів належать рецесивні гени гемофілії, дальтонізму, домінантний ген гіпофосфатемії (вітамін D-резистентний рахіт).
Другу групу складає невелике число генів, теж повністю зчеплених зі статтю, але розташованих у негомологічній ділянці Y-хромосоми. Вони передаються від батька до всіх його синів, бо Y-хромосому син одержує лише від батька (перетинки між пальцями, "волосаті вуха"). Ознаки, гени яких розміщені в негомологічній ділянці Y-хромосоми, називаються голандричними.
Третю групу складають гени, неповно (частково) зчеплені зі статтю. Вони розташовані в гомологічних ділянках X- і Y-хромосом, можуть передаватися як з Х-, так і з Y-хромосомою, і переходити з однієї в іншу при кросинговері.
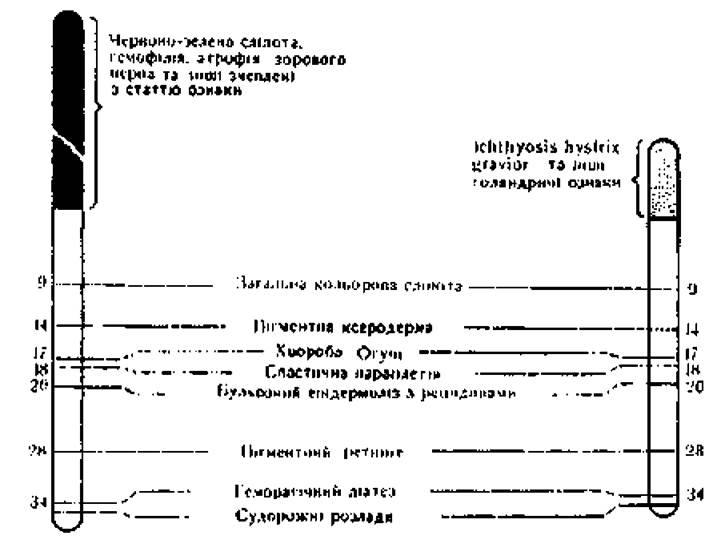
Рис. 21. Найімовірніше розташування в гомологічних ділянках X- та Y-хромосом генів, неповністю зчеплених зі статтю.
У людини ознаки, які успадковуються через Y -хромосому, можуть бути тільки у особин чоловічої статі, а успадковані через Х-хромосому - у особин як однієї, так і іншої статті. Особина жіночої статі може бути як гомо- так і гетерозиготною за генами, які локалізовані у Х-хромосомі, а рецесивні алелі генів у нього проявляються тільки у гомозиготному стані. Оскільки у осіб чоловічої статі тільки одна Х-хромосома, всі локалізовані у ній гени, навіть рецесивні, проявляються у фенотипі. Такий організм називають гемізиготним.
Гени, які визначають ознаки статі, знаходяться не тільки в статевих хромосомах. Гени ознак, обмежених статтю, можуть бути розташовані як в автосомах, так і в статевих хромосомах обох статей, але проявляються лише в однієї з них. Наприклад, баритон і бас - проявляються лише в чоловіків; молочність і жирність молока-лише в корови; яйценоскість і розмір яєць - лише в курки.
Ознаки, залежні від статі - це ознаки детерміновані аутосомними генами у чоловіків і жінок, але домінування цих ознак залежить від статевих гормонів. Залежними від статі ознаками є успадкування лисості у чоловіків і жінок. Ген лисості — домінантний. Його рецесивна алель визначає нормальне волосся. Домінантний ген в гомозиготному стані (ВВ) проявляється майже однаково у чоловіків і жінок — чоловіки будуть лисі. Жінки з рідким волоссям. У рецесивних гомозигот (вв) також однакові прояви ознак в обох статей. Якщо ген лисості знаходиться в гетерозиготному стані (Вв) у чоловіків — вони усі лисі; у гетерозиготних жінок дані ознаки не проявляться, вони будуть мати нормальне волосся. Стать впливає у людини і на таку ознаку, як подагра. При подагрі в суглобах відкладаються солі сечової кислоти, викликаючи сильний біль у людини. Ген, який відповідає за це захворювання, проявляється в присутності чоловічих статевих гормонів значно сильніше, ніж в присутності жіночих.
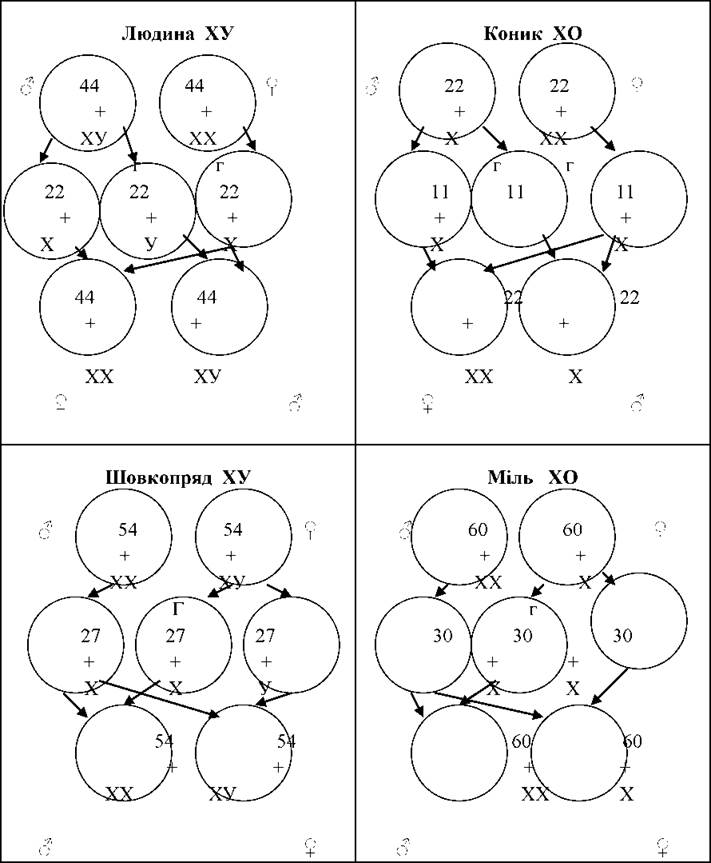
Рис. 22. Схема різних типів хромосомного визначення статі.
Стать - це сукупність властивостей організму, який забезпечує функцію відтворення потомства на основі передачі генетичної інформації. Розрізняють первинні і вторинні статеві ознаки. До, спарювання і запліднення, але відіграють певну первинних статевих ознак відносять всі ознаки організму, які забезпечують утворення гамет, злиття їх в процесі запліднення, а також відмінності в будові внутрішніх і зовнішніх органів розмноження. До вторинних статевих ознак відносяться ознаки, які безпосередньо не впливають на процес гаметогенезароль при статевому розмноженні (грудні залози у ссавців, оперення у птахів). Розвиток вторинних статевих ознак контролюється в системі всього організму гормональною діяльністю, і як наслідок, безпосередньо пов’язаний з функцією первинних статевих залоз.
Фізіологічна теорія визначення статі говорить про те, що організм є генетично бісексуальним і в генотипі знаходяться гени обох статей, але співвідношення цих генів, можливо, і характер їх дій у чоловічих і жіночих особин різний. Чоловіки мають гени чоловічої статі не тільки в Y-хромосомі, в невеликій кількості вони містяться і в Х- хромосомі а також в аутосомах. Жінки, як і чоловіки, також мають гени як жіночої, так і в невеликій кількості чоловічої статі. Процес диференціювання статі зумовлений статевими гормонами, які виділяються не тільки ендокринними залозами, але й гормонами зачаткової статевої залози: чоловічий статевий гормон зачаткової статевої залози — медулларин, жіночий гормон -- кортикальний гормон. Рівень гормональної секреції контролюється балансом генів. Перевага генів чоловічої статі призводить до підвищення секреції чоловічих статевих гормонів і до формування чоловічої статі. Перевага жіночих статевих гормонів веде до розвитку особини жіночої статі. Зміна активностей гормональної секреції однієї статі, або другої призводить до розвитку інтерсексуальних форм, яких називають гермафродитами.