Фізіологія людини - Вільям Ф. Ґанонґ 2002
Ендокринна система, метаболізм і репродуктивна функція
Енергетичний баланс, метаболізм і живлення
Метаболізм білків
Білки
Білки складаються з великої кількості амінокислот (рис. 17-16), з’єднаних у ланцюги пептидними зв’язками, які сполучають аміногрупу однієї кислоти з карбоксильною групою іншої. Окрім цього, деякі білки містять вуглеводи (глікопротеїни) і ліпіди (ліпопротеїни). Менші ланцюги амінокислот називають пептидами або поліпептидами. Межі між пептидами, поліпептидами та білками визначені не чітко. У цій книзі ланцюги завдовжки 2-10 амінокислотних залишків названо пептидами, ланцюги, що містять їх понад 10, але менше 100, - поліпептидами, а ланцюги, що містять 100 і більше амінокислотних залишків, - білками. Термін олігопептид, який деколи застосовують для позначення малих пептидів, ми не використовуємо. Порядок розташування амінокислот у пептидному ланцюгу називають первинною структурою білка. Ланцюги, зігнуті й скручені складним способом, і термін вторинна структура білка стосується його просторової організації, утвореної згинанням і скручуванням. Типовою вторинною структурою є правильна спіраль із трьома, сімома амінокислотними залишками у витку (а-спіраль). Інша типова вторинна структура - ß-складчастий шар. Антипаралельний ß-складчастий шар утворюється, коли розтягнутий поліпептидний ланцюг згортається з утворенням водневих зв’язків між пептидними зв’язками сусідніх ланцюгів. Крім того, паралельні ß-складчасті шари виникають між поліпептидними ланцюгами. В так званій стрічковій моделі білка (див. рис. 27-8) а-спіралі показані у вигляді витків, а ß-складчасті шари - у вигляді паралельних стрілок.
Третинна структура білка - це упакування згорнутих ланцюгів у шари, кристали чи волокна. Термін четвертинна структура використовують для позначення розташування субодиниць, із яких складено багато білкових молекул (наприклад, гемоглобін; див. Розділ 27).
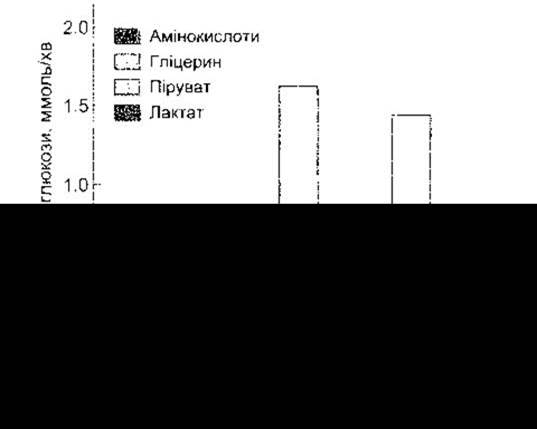
Рис. 17-15. Вихід з печінки глюкози, утвореної внаслідок глікогенолізу (незабарвлена частина стовпця), і глюкози, утвореної завдяки, вірогідно, глюконеогенезу (обмежена фігурною дужкою частина стовпця). Показано внесок у глюконеогенез різних попередників глюкози (відтворено за дозволом з Felig Р, Wahren J: Fuel homeostasis in exercise. N Engl J Med 1975;293:1078).
Амінокислоти
Амінокислоти, що містяться в білках, наведені в табл. 17-2. Їх позначено три- чи однобуквеними скороченнями. Багато інших важливих амінокислот, таких як орнітин, 5- гідрокситриптофан, L-ДОФА, таурин та тироксин (Т4), є в організмі, однак їх не виявлено в білках. У вищих тварин природними формами амінокислот є L-ізомери. Зокрема, L-ізомери гормонів, наприклад, тироксину, набагато активніші, ніж D-ізомери. Амінокислоти мають основну, нейтральну чи кислу реакцію залежно від співвідношення вільних кислотних (-СООН) та основних (-NH2) груп у молекулі.
Деякі з амінокислот є незамінними амінокислотами, тобто їх потрібно отримувати з їжею, тоді як інші можуть синтезуватись in vivo у кількостях, достатніх для задоволення метаболічних потреб (див. нижче).
Пул амінокислот
Незважаючи на те, що в шлунково-кишковому тракті відбувається всмоктування невеликої кількості білків та деяких пептидів, більшість спожитих білків перетравлюються і їхні складові амінокислоти всмоктуються. Білки самого організму постійно розщеплюються до амінокислот і ресинтезуються. Обіг ендогенних білків у середньому становить 80-100 г/д і є найбільшим у слизовій оболонці кишки та майже нульовим у колагені. Амінокислоти, утворенні під час ендогенного розпаду білків, ідентичні до амінокислот білків їжі. Разом вони формують пул амінокислот, що забезпечує потреби організму (рис. 17- 17). У нирках відбувається реабсорбція більшості відфільтрованих амінокислот. Під час росту рівновага між амінокислотами та білками організму зсувається в бік білків так, що синтез перевищує розпад. За будь-якого віку частина білків утрачається у вигляді волосся, у жінок невеликі кількості білків відходять під час місячних. Деякі малі білки виходять із сечею, а у випорожненнях наявні нереадсорбовані білки травних секретів. Утрати поповнюються шляхом синтезування з пулу амінокислот.
Таблиця 17-2. Амінокислоти, що містяться в білках

* У дужках наведено загальноприйняті три- і однобуквені скорочення амінокислот.
** Жирним шрифтом виділені незамінні амінокислоти.
1 Нема тРНК для цих чотирьох амінокислот; вони утворені посттрансляційною модифікацією відповідних немодифікованих амінокислот у пептидному зв’язку. Є тРНК для селеноцистеїну та інших 20 амінокислот, і вони входять у білки під прямим генетичним контролем.
2 Аргінін і гістидин іноді називають напівнезамінними - вони не є необхідними для підтримання азотного балансу, однак потрібні для нормального росту.
3 Селеноцистеїн є рідкісною амінокислотою, у якій сірка цистеїну замінена селеном. Кодон LIGA зазвичай є стоп-кодоном, проте в деяких випадках він кодує селеноцистеїн.
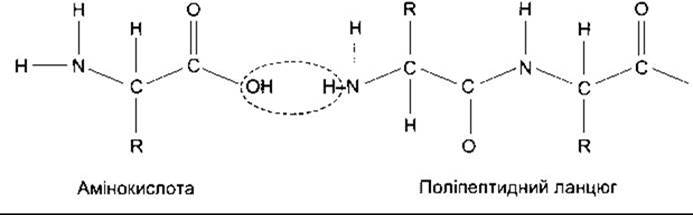
Рис. 17-16. Структура амінокислот і утворення пептидних зв’язків. Штрихова лінія відображає утворення пептидних зв’язків з виділенням Н2О; R - амінокислотний залишок; наприклад, у гліцині R = Н, у глутаматі R = -(СН2)2-СОО-.
Специфічні метаболічні функції амінокислот
Тиреоїдні гормони, катехоламіни, гістамін, серотонін, мелатонін та проміжні сполуки в циклі сечовини утворюються з деяких амінокислот. Метіонін та цистеїн постачають сірку, що міститься в білках, КоА, таурині та інших біологічно важливих сполуках. Метіонін перетворюється в S-аденозилметіонін - активний метилювальний агент у синтезі таких сполук, як адреналін. Він є головним донором біологічно лабільних метальних груп, однак метальні групи можуть також надходити з похідними мурашиної кислоти, приєднаними до похідних фолієвої кислоти, якщо в харчовому раціоні є достатня кількість фолієвої кислоти та ціанокобаламіну.
Сульфати сечі
Окиснення цистеїну - головне джерело сульфатів у сечі. Більшість екскрецій сечі є у вигляді сульфатів (SО42 ), які супроводжують відповідні кількості катіонів (Na+, К+, NH4+, чи Н+). Ефірні сульфати в сечі - це органічні естери сульфокислот (R-О-SО3H), що утворюються в печінці з ендогенних та екзогенних фенолів, у тім числі естрогенів та інших стероїдів, індолів та ліків.
Дезамінування, амінування і трансамінування
Взаємоперетворення між амінокислотами та продуктами катаболізму вуглеводів і жирів на рівні загального метаболічного пулу та циклу лимонної кислоти охоплюють перенесення, відщеплення та утворення аміногруп.
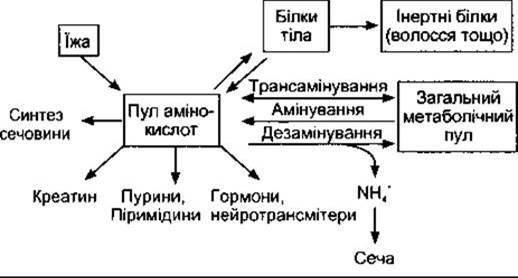
Рис. 17-17. Метаболізм амінокислот.
Реакції трансамінування, перетворення однієї амінокислоти у відповідну кетокислоту з одночасним перетворенням іншої кетокислоти в амінокислоту:
![]()
відбуваються у багатьох тканинах. Трансамінази, що беруть участь у цих процесах, наявні також у кровообігу. Якщо виникає ушкодження багатьох активних клітин як наслідок патологічного процесу, то рівень сироваткових трансаміназ зростає. Прикладом є підвищення рівня аспартат-амінотрансферази (ACT) у плазмі крові у разі інфаркту міокарда.
Окиснювальне дезамінування амінокислот відбувається в печінці. Імінокислота утворюється шляхом дегідрогенації, далі реакція гідролізу приводить до відповідної кетокислоти з утворенням NH4+:
![]()
Катіон NH4+ перебуває в рівновазі з NH3. Амінокислота може також приєднувати NH4+, утворюючи відповідний амід. Прикладом такої реакції є приєднання NH4+ до глутамату, яке фіксують у мозку (рис. 17-18). Зворотна реакція відбувається в нирках з утворенням NH4+ і його екскрецією із сечею (див. Розділ 38).
Взаємоперетворення між пулом амінокислот і загальним метаболічним пулом відображені на рис. 17-19. Лейцин, ізолейцин, фенілаланін та тирозин називають кето-генними, оскільки вони перетворюються в ацетоацетат (кетонові тіла, див. нижче). Аланін та багато інших амінокислот є глюкогенними, бо вони дають початок сполукам, які можуть легко перетворитися в глюкозу.
Біосинтез сечовини
Більшість NH4+, утвореного деамінуванням амінокислот у печінці, перетворюється в сечовину. Катіон NH4+ утворює карбамоїлфосфат, який у мітохондріях переноситься на орнітин, утворюючи цитрулін. Ензимом, що каталізує процес, є карбамоїлтрансфераза. Цитрулін перетворюється в аргінін, від якого відщеплюється сечовина, і регенерується орнітин (цикл сечовини; рис. 17-20). Більшість сечовини утворюється в печінці, і в разі серйозних хвороб печінки азот сечовини крові (АСК) знижується, а рівень NH3 у крові підвищується. Спадковий дефіцит орнітинкарбомоїлтрансферази також може призвести до інтоксикації NH3 навіть в осіб, що є гетерозиготними за цією ознакою.
Креатин і креатинін
Креатин синтезується в печінці з метіоніну, гліцину й аргініну. У скелетних м’язах він фосфорилюється з утворенням фосфокреатину (рис. 17-21), який є важливим резервом енергії для синтезу АТФ (див. Розділ 3). АТФ, що виникає під час гліколізу та окисного фосфорилювання, реагує з креатином, завдяки чому утворюється АДФ та фосфокреатин. Під час фізичних навантажень реакція відбувається у зворотному напрямі, забезпечуючи постачання АТФ, який є безпосереднім джерелом енергії для скорочення м’язів. Деякі атлети споживають креатин як харчовий додаток і стверджують, що він підвищує їхні результати в спринті та інших формах короткотривалих навантажень.
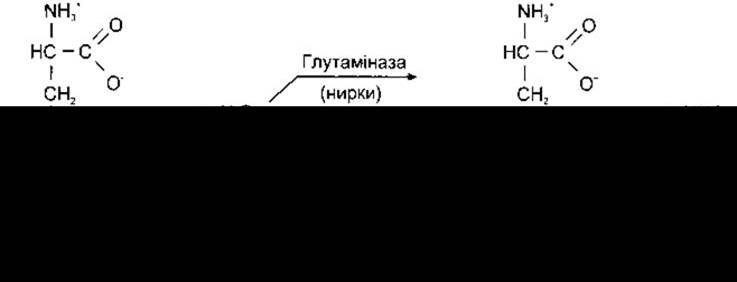
Рис. 17-18. Вивільнення та приєднання NH4+ під час взаємоперетворення глутаміну і глутамату. Катіон NH4+ перебуває в стані рівноваги з NH3. Рівновага реакції зміщується правобіч у нирках, і NH3 секретується в сечу. Рівновага реакції' зміщується лівобіч у головному мозку, видаляючи токсичний для нервових клітин NH3.
Креатинін у сечі утворюється з фосфокреатину. Креатин прямо не перетворюється в креатинін. Рівень екскреції креатиніну порівняно сталий протягом тривалого часу. Під час дослідження метаболізму часто вимірюють кількість креатиніну в сечі для перевірки точності. Визначають середнє денне виділення креатиніну і значення денного утворення інших сполук співвідносять до певного рівня виведеного креатиніну.
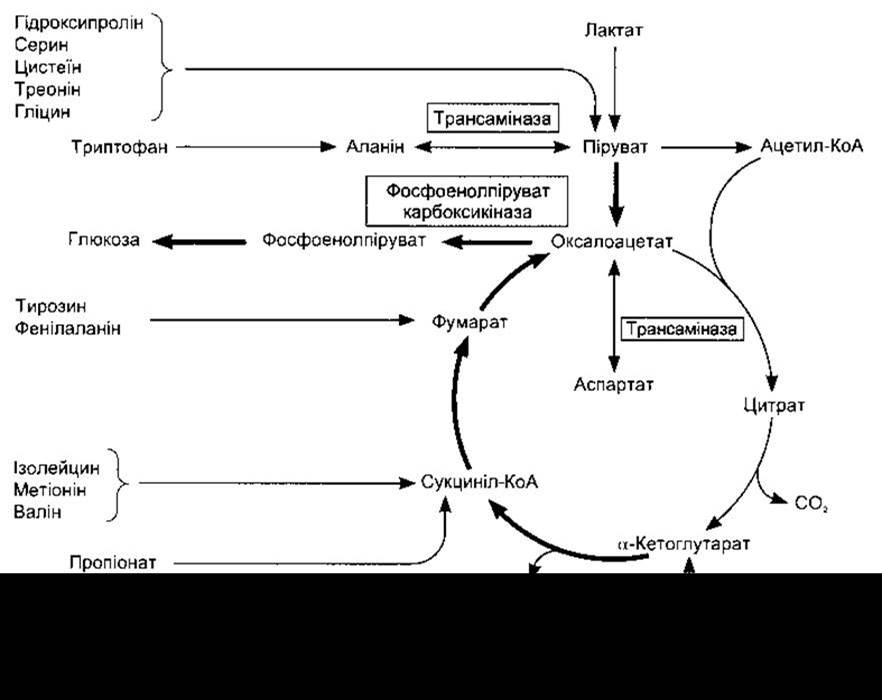
Рис. 17-19. Участь циклу лимонної кислоти в трансамінуванні та глюконеогенезі. Виділені стрілки позначають головний шлях глюконеогенезу (відтворено за дозволом з Murray RK et al: Harper’s Biochemistry, 25th ed. McGraw-Hill, 2000).
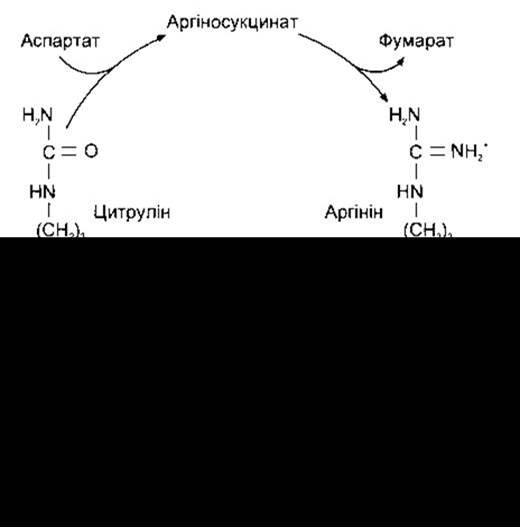
Рис. 17-20. Цикл сечовини.
Креатинурія інколи простежується в дітей, жінок під час та після вагітності й інколи в невагітних жінок. У нормі в сечі чоловіків є дуже мало креатину або ж узагалі нема, проте за будь-якого стану, пов’язаного з інтенсивним м’язовим розпадом його рівень помітно збільшується. Креатинурія виникає внаслідок голодування, тиреотоксикозу, некомпенсованого цукрового діабету та різноманітних первинних і вторинних хвороб м’язів (міопатіях).
Пурини та піримідини
Фізіологічно важливі пурини та піримідини показані нарис. 17-22. Нуклеозиди - пурини чи піримідини, з’єднані з рибозою, - є компонентами не тільки різноманітних коензимів та пов’язаних із ними сполук (НАД+, НАДФ+, АТФ, УДФГ тощо), а також РНК та ДНК (табл. 17-3). Структура та функції ДНК і РНК, а також їхня роль у синтезуванні білків розглянуті у Розділі 1.
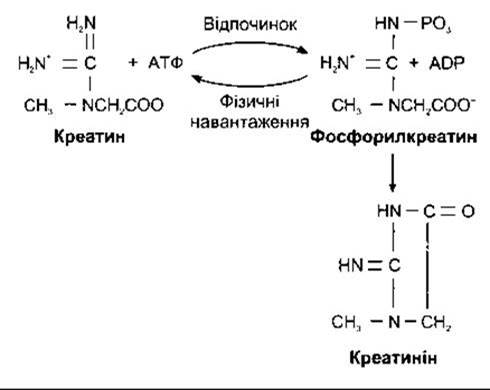
Рис. 17-21. Креатин, фосфорилкреатин та креатинін.
Нуклеїнові кислоти їжі розщеплюються (перетравлюються), а їхні складові пурини і піримідини абсорбуються, проте більшість пуринів і піримідинів синтезується з амінокислот головно в печінці, з яких далі синтезуються нуклеотиди та РНК і ДНК. РНК перебуває в динамічній рівновазі з пулом амінокислот, однак ДНК, будучи сформованою, метаболічно стабільна протягом життя.
Пурини та піримідини, вивільнені внаслідок розпаду нуклеотидів, можуть знову бути використані або катаболізовані. Лише невелика їхня частина виводиться в незміненому стані із сечею. Піримідини катаболізуються до С02 і NH3, а пурини перетворюються в сечову кислоту.
Розпад білків
Як і синтез білків, їхній розпад є ретельно регульованим складним процесом. Сполучення білків із 74-амінокислот- ним поліпептидом убіквітином є позначкою для білків, призначених для розпаду. Цей поліпептид високо консервативний і наявний в усіх клітинах - від бактерій до людини. Убіквітинація цитоплазматичних білків, у тім числі інтегральних білків ендоплазматичної сітки, маркує білки для розпаду в багатосубодиничних протеолітичних частинках - 26S протеасомах, а убіквітинація мембранних білків, таких як рецептори гормону росту, маркує їх для розпаду в лізосомах. Зафіксовано беззаперечний баланс між інтенсивністю синтезу білків та їхнього розпаду, отже, приєднання убіквітину має важливе значення в клітинній біології, наприклад, у регулюванні клітинного циклу (див. Розділ 1). Рівні метаболізму окремих білків відрізняються, а тому в організмі є механізм, за допомогою якого аномальні білки розпізнаються й руйнуються швидше, ніж нормальні складові. Наприклад, аномальні гемоглобіни швидко катаболізуються в осіб зі спадковими гемоглобінопатіями (див. Розділ 27). Розпад білків зменшується у гіпертрофованих тренованих скелетних м’язах і збільшується під час атрофії у денервованому м’язі чи скелетному м’язі, що з якихось причин не працює. Окрім того, він є чинником у визначенні розміру органа (наприклад, рівень розпаду печінкових білків помітно зменшується під час компенсаторної гіпертрофії, яка настає після часткової гепатектомії).
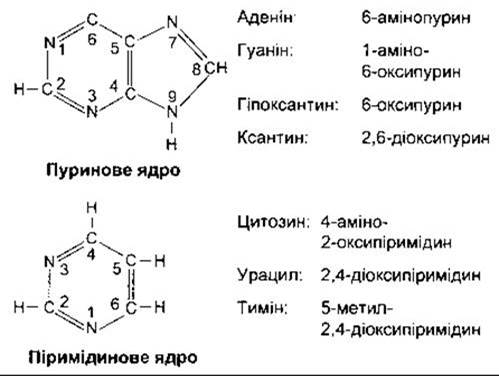
Рис. 17-22. Головні фізіологічно важливі пурини та піримідини. Оксипурини та оксипіримідини можуть утворювати енольні похідні (гідроксипурини та гідроксипіримідини) шляхом переміщення атома водню до кисневмісного замісника.
Таблиця 17-3. Пурино- і піримідиновмісні сполуки
Тип сполуки |
Компоненти |
Нуклеозид |
Пурин чи піримідин плюс рибоза чи 2-дезоксирибоза |
Нуклеотид (мононуклеотид) |
Нуклеозид плюс залишок фосфорної кислоти |
Нуклеїнова кислота |
Багато нуклеотидів, що утворюють дволанцюгові структури чи два полінуклеотидні ланцюги |
Нуклеопротеін |
Нуклеїнова кислота плюс один чи більше простих основних білків |
Рибозовмісні |
Рибонуклеїнова кислота |
2-Дезоксирибозовмісні |
Дезоксирибонуклеїнова кислота |
Сечова кислота
Сечова кислота утворюється внаслідок розпаду пуринів і прямого синтезування з 5-фосфорибозилпірофосфату (5- ФРПФ) та глутаміну (рис. 17-23). У людей вона виводиться з сечею, а в деяких тварин перед екскрецією окиснюється до алантоїну. Нормальний рівень сечової кислоти в крові людини становить близько 4 мг/дл (0,24 ммоль/л). У нирках відбувається її фільтрування, реабсорбування й виведення. Зазвичай 98% відфільтрованої сечової кислоти реадсорбується, а решта 2% відповідають близько 20% кількості сечової кислоти, яка виводиться. Інші 80% надходять завдяки канальцевій секреції. Виведення сечової кислоти в разі безпуринової дієти становить 0,5 г/добу, а у випадку нормального харчування - близько 1 г/добу.
Первинна та вторинна подагра
Подагра - це хвороба з повторюваними приступами артриту, уратними відкладеннями в суглобах, нирках та інших тканинах, а також збільшеним рівнем сечової кислоти в крові та сечі. Спочатку зазнає враження плеснофаланговий суглоб великого пальця ноги. Є дві форми первинної подагри: у разі одної збільшується утворення сечової кислоти внаслідок різних ензимних аномалій; у випадку іншої вибірково порушується канальцеве транспортування сечової кислоти в нирках. У разі вторинної подагри рівень сечової кислоти в рідинах організму збільшується внаслідок зменшення екскреції або посиленого її утворення, яке є вторинним щодо якоїсь хвороби. Наприклад, екскреція зменшується в пацієнтів, які вживають триазидні сечогінні засоби (див. Розділ 38), та в осіб із хворобами нирок. Утворення сечової кислоти збільшується в разі лейкемії та пневмонії внаслідок активного руйнування лейкоцитів.
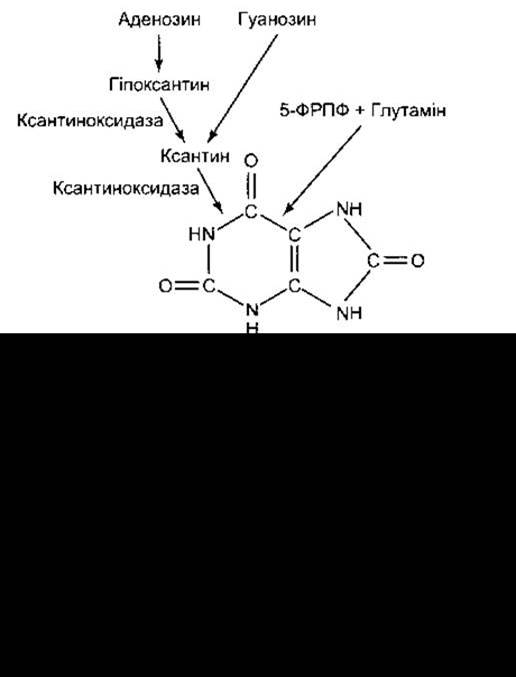
Рис. 17-23. Синтезування та розпад сечової кислоти. Аденозин переходить у гіпоксантин, який потім перетворюється в ксантин, а ксантин - у сечову кислоту. Дві останні реакції каталізує ксантиноксидаза. Гуанозин прямо перетворюється в ксантин. Ксантиноксидазу інгібує алопуринол - один з препаратів, який застосовують для лікування подагри.
Лікування подагри полягає у зниженні симптоматики гострого артриту за допомогою таких ліків, як колхіцин чи нестероїдні протизапальні препарати, та зменшенні рівня сечової кислоти в крові. Колхіцин не впливає на метаболізм сечової кислоти, він, очевидно, зменшує приступи подагри, інгібуючи фагоцитоз кристалів сечової кислоти лейкоцитами, тобто певним чином зумовлюючи суглобні симптоми. Фенілбутазон та пробенецид інгібують реабсорбцію сечової кислоти в ниркових канальцях. Алопуринол, що інгібує ксантиназу (див. рис. 17-23), є одним з препаратів, який використовують із метою зменшити утворення сечової кислоти.
Азотний баланс
Для заміни втрачених білків і амінокислот потрібно щоденно споживати певну кількість білка. Це стосується не самих білків, а їхніх складових - амінокислот, які можна споживати в чистому вигляді. Втрата білків та їхніх похідних із випорожненнями зазвичай дуже мала. Тому кількість азоту в сечі є надійним показником кількості незворотного розпаду білків і амінокислот. Якщо кількість азоту в сечі дорівнює вмісту азоту в білку їжі, то кажуть, що в особи настав азотний баланс. Зі зростанням споживання білків більша кількість амінокислот деамінується, екскреція сечовини посилюється, і азотний баланс відновлюється. Незважаючи на це, у випадках, коли підвищена секреція катаболічних гормонів кори надниркових залоз чи зменшена секреція інсуліну, та під час голодування чи вимушеної іммобілізації втрати азоту перевищують його надходження, і азотний баланс стає негативним. Під час росту, одужання від хвороби чи введення анаболічних стероїдів, таких як тестостерон, надходження азоту перевищує його екскрецію, і азотний баланс стає позитивним.
У випадку нестачі однієї з незамінних амінокислот, що потрібна для синтезування білків, білок не синтезується. Інші амінокислоти, які повинні бути у складі білка, деамінують, як і інші надлишкові амінокислоти, а їхній азот виділяється у вигляді сечовини. Це є причиною того, що азотний баланс стає негативним, якщо одну з незамінних амінокислот вилучають із харчового раціону.
Відповідь на голодування
Якщо особа споживає їжу, яка містить мало білків, однак є достатньо калорійною, то екскреція сечовини та сульфо-ефірів знижується; екскреція сечової кислоти зменшується на 50%, а екскреція креатину не змінюється. Внаслідок цього креатин та близько половини сечової кислоти в сечі утворюються тільки завдяки процесу «зношення», на який не впливає кількість спожитого білка. Загальна екскреція азоту менше 3,6 г/добу під час білкового голодування, навіть коли їжа достатньо калорійна, оскільки негативним є азотний баланс, спричинений дефіцитом незамінних амінокислот.
У випадку недостатньо калорійного харчування екскреція азоту в сечовині в середньому становить 10 г/добу, тому що білки розщеплюються для отримання енергії. Невеликі кількості глюкози протидіють цьому катаболізму до помітного рівня (глюкозний ефект білкового резервування). Такий ефект виникає головно внаслідок того, що глюкоза збільшує секрецію інсуліну, який, відповідно, гальмує розпад білків у м’язах, а також після введення порівняно невеликих кількостей амінокислот.
Жири теж сприяють затриманню азоту. Під час тривалого голодування кетокислоти, утворені з жирів (див. нижче), використовуються у мозку та інших тканинах. Ці речовини є кофакторами метаболізму в м’язах разом із трьома амінокислотами, що мають розгалужений ланцюг: лейцином, ізолейцином та валіном. Під час утилізації жиропохідних кетокислот ці амінокислоти, очевидно, нагромаджуються. Введення безазотних аналогів названих амінокислот зумовлює білкове резервування та зменшує утворення сечовини й аміаку в пацієнтів з ушкодженнями нирок і печінки.
Більшість білків, розпад яких відбувається під час загального голодування, надходить з печінки, селезінки та м’язів і, порівняно мало, - із мозку та серця. Після вичерпання запасів глікогену в печінці рівень глюкози в крові дещо знижується (див. вище), однак завдяки глюконеогенезу він є вищим від того рівня, що спричинює гіпоглікемічні симптоми. Простежується кетонемія, нейтральні жири швидко катаболізують. Коли запаси жиру вичерпані, то катаболізм білків ще більше посилюється і швидко настає смерть. Середня людина масою 70 кг має 0,1 кг глікогену в печінці, 0,4 кг глікогену в м’язах та 12 кг жиру. Цих запасів глікогену достатньо для одноденного голодування. В госпіталізованих опасистих пацієнтів, яким дають лише воду та вітаміни, простежується втрата маси в межах 1 кг/добу протягом перших 10 днів. Потім втрата маси зменшується й стабілізується на значенні близько 0,3 кг за добу. Упродовж деякого часу пацієнти почуваються досить добре, проте інколи трапляються ускладнення у вигляді постуральної гіпотензії та подагричного артриту. У людини середній час від початку голодування до смерті становить 60 днів.