Фізіологія людини - Вільям Ф. Ґанонґ 2002
Ендокринна система, метаболізм і репродуктивна функція
Щитоподібна залоза
Утворення та секреція тиреоїдних гормонів
Хімія
Головними гормонами, які секретує щитоподібна залоза, є трийодотиронін (T3) та тироксин (Т4). Трийодотиронін також утворюється в периферійних тканинах шляхом дейодування Т4 (див. нижче). Обидва гормони є йодовмісними амінокислотами (рис. 18-4). У венозній крові щитоподібної залози виявлено невеликі кількості оберненого трийодотироніну (3,3',5'-трийодотироніну, ОТ3) та інших сполук. Трийодотиронін активніший, ніж Т4, тоді як ОТ3 неактивний. Природні форми Т4 і його похідні з асиметричним атомом вуглецю є L-ізомерами. Водночас D-тироксин становить лише невелику частину активності L-форми.
Тиреоглобулін
Гормони Т3 та Т4 синтезуються в колоїді шляхом йодування, а також конденсації молекул тирозину, з’єднаних пептидним зв’язком із тиреоглобуліном. Цей глікопротеїн складається з двох субодиниць і має молекулярну масу 660 000. Близько 10% його маси становлять вуглеводи. Він також містить 123 залишки тирозину, однак лише від чотирьох до восьми з них зазвичай входять у тиреоїдні гормони. Синтезування тиреоглобуліну відбувається в фолікулярних епітеліальних клітинах щитоподібної залози, а секретування в колоїд шляхом екзоцитозу - у гранулах, що також містять тиреопероксидазу (див. нижче). Гормони зв’язані з тиреоглобуліном аж до секреції. Після секреції колоїд поглинають клітини щитоподібної залози, і пептидні зв’язки гідролізуються, в цьому разі вільні Т3 і Т4 виходять у капіляри. Отже, фолікулярні клітини щитоподібної залози виконують три функції: накопичують та транспортують йод; синтезують тиреоглобулін і секретують його в колоїд; відщеплюють тиреоїдні гормони від тиреоглобуліну й секретують їх у кров.
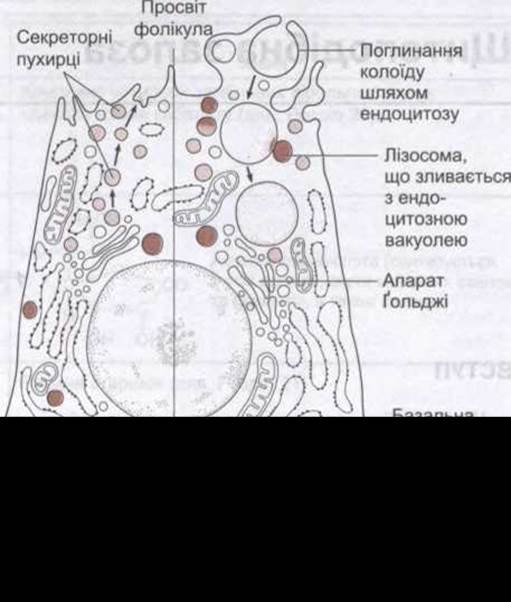
Рис. 18-3. Клітина щитоподібної залози. Ліворуч: нормальна. Праворуч: після помітного стимулювання ТТГ. Стрілки ліворуч показують секрецію тиреоглобуліну в колоїд. Праворуч відображено ендоцитоз колоїду та злиття вакуолі, що містить колоїд, з лізосомою. Клітини примикають до капілярів з отворами (фенестраціями) ендотеліальної стінки.
Тиреоглобулін надходить у кров разом із колоїдом. Нормальна концентрація тиреоглобуліну в плазмі становить близько 6 нг/мл, і цей рівень підвищується в разі гіпертиреоїдизму та деяких видів раку щитоподібної залози. Проте функція тиреоглобуліну, що циркулює в крові, якщо така є, невідома.
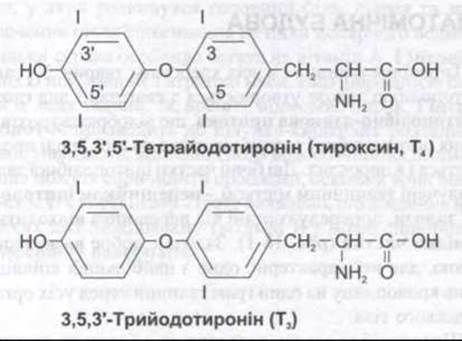
Рис. 18-4. Тиреоїдні гормони. Числа в кільцях у формулі Т4 відображають місце розташування йоду в молекулі.
Метаболізм йоду
Йод є вихідним матеріалом для синтезування тиреоїдних гормонів. Спожитий йод перетворюється в йодид і абсорбується. Частка абсорбованого І показана на рис. 18-5. Мінімальне добове поглинання йоду, яке підтримує нормальну діяльність щитоподібної залози, становить 150 мкг у дорослих (див. табл. 17-7), проте у розвинутих країнах середнє поглинання йоду з їжею дорівнює 500 мкг/добу. Нормальний рівень І у плазмі - близько 0,3 мкг/дл, а розподіляється він у об’ємі 25 л (35% від маси тіла). Головними органами, що накопичують І , є щитоподібна залоза, яка використовує його для синтезування тиреоїдних гормонів, та нирки, які екскретують його із сечею. За нормальних рівнів синтезу та секреції тиреоїдних гормонів у щитоподібну залозу надходить 120 мкг/добу. Щитоподібна залоза секретує 80 мкг/добу у вигляді йоду в Т3 і Т4. Сорок міліграмів І у день дифундує в ПКР. Секретовані Т3 і Т4 метаболізуються в печінці та інших тканинах зі щоденним вивільненням 60 мкг І у ПКР. Деякі похідні тиреоїдних гормонів екскретують у жовчі, і незначна частина йоду в них реабсорбується (ентерогепатична циркуляція), однак сумарна втрата І із випорожненнями становить близько 20 мкг/добу. Отже, загальне надходження І в ПКР - 500 + 40 + 60 = 600 мкг/добу; 20% цього І надходить у щитоподібну залозу, тоді як 80% екскретує з сечею.
Концентрування йодиду
Щитоподібна залоза концентрує йодид шляхом активного його транспортування з кровообігу в колоїд. Цей транспортний механізм часто називають механізмом концентрування йодиду, або йодидною помпою. Помпа є прикладом вторинного активного транспортування (див. Розділ 1); Na+ та І котранспортують у клітини щитоподібної залози, a Na+ накопичує в інтерстиції 1Ча+-К+-АТФ-аза. Клоновано котранспортер Na+-I , він має 12 трансмембранних доменів з аміно- та карбоксильними кінцями всередині клітини.
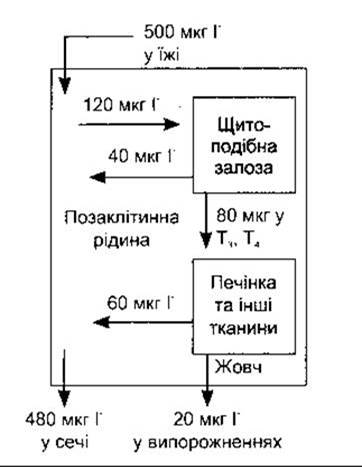
Рис. 18-5. Метаболізм йоду.
Клітина щитоподібної залози приблизно на 50 мВ негативніша порівняно з інтерстиційною ділянкою колоїду; тобто вона має потенціал спокою мембрани -50 мВ. Йодид накопичується в клітині проти цього електричного градієнта, а потім дифундує за градієнтом у колоїд. Поглинання йоду можна вивчати шляхом уведення радіоактивного йоду у відстежувальних дозах, що становлять малі кількості, які не можуть суттєво підвищити рівень йодиду в організмі. В залозі йодид швидко окиснюється і зв’язується з тирозином. Незважаючи на це, співвідношення вільного йодиду тиреоїду до йодиду сироватки плазми (відношення Т/С) за нормальних умов більше від 1. Якщо зв’язування із залишками тирозину блокують антитиреоїдні препарати, такі як пропілтіоурацил (див. нижче), то йодид накопичується в щитоподібній залозі, а співвідношення Т/С помітно збільшується. Перхлорат та багато інших аніонів зменшують транспортування йодиду шляхом конкурентного інгібування.
Варто зазначити, що реагування щитоподібної залози на йодид є унікальним; йодид потрібний для нормальної діяльності щитоподібної залози (це детально описано нижче), однак і його нестача, і надлишок гальмують функції цього органа.
Слинні залози, слизова оболонка кишки, плацента, війчасте тіло ока, судинне сплетення та молочні залози також транспортують йодид проти градієнта концентрації, проте на їхнє поглинання не впливає ТТГ. Молочні залози теж зв’язують йод; дийодотирозин утворюється в тканині молочних залоз, а Т3 і Т4 - ні. Фізіологічна важливість усіх таких позащитоподібних йодидоконцентрувальних механізмів остаточно не з'ясована.
Синтез тиреоїдних гормонів
У щитоподібній залозі йодид окиснюється до йоду й за лічені секунди зв’язується в трьох положеннях молекул тирозину, приєднаного до тиреоглобуліну (рис. 18-6). Ензимом, відповідальним за окиснення та зв’язування йоду, є тиреопероксидаза, а також пероксид водню, що приймає електрони. Після цього утворений монойодотирозин (MIT) йодується в положенні 5 з утворенням дийодотирозину (ДІТ). Дві молекули ДІТ зазнають окиснювальної конденсації з утворенням Т4 та відщепленням бічного ланцюга аланіну від молекули, яка утворює зовнішнє кільце. Є дві теорії того, як відбувається реакція сполучення. За однією сполучення відбувається між двома молекулами ДІТ, зв’язаними з тиреоглобуліном (внутрішньомолекулярне сполучення); за іншою ДІТ, який утворює зовнішнє кільце, спершу від’єднується від тиреоглобуліну (міжмолекулярне сполучення). У будь-якому випадку, тиропероксидаза задіяна, крім йодування, також у сполученні. Ймовірно, Т3 утворюється шляхом конденсації MIT та ДІТ. Унаслідок конденсації ДІТ і MIT, мабуть, утворюються також невеликі кількості ОТ3. У нормальній щитоподібній залозі людини середній розподіл йодованих сполук становить 23% MIT, 33 - ДІТ, 35 - Т4 та 7% Т3, і лише сліди припадають на ОТ3 та інші сполуки.
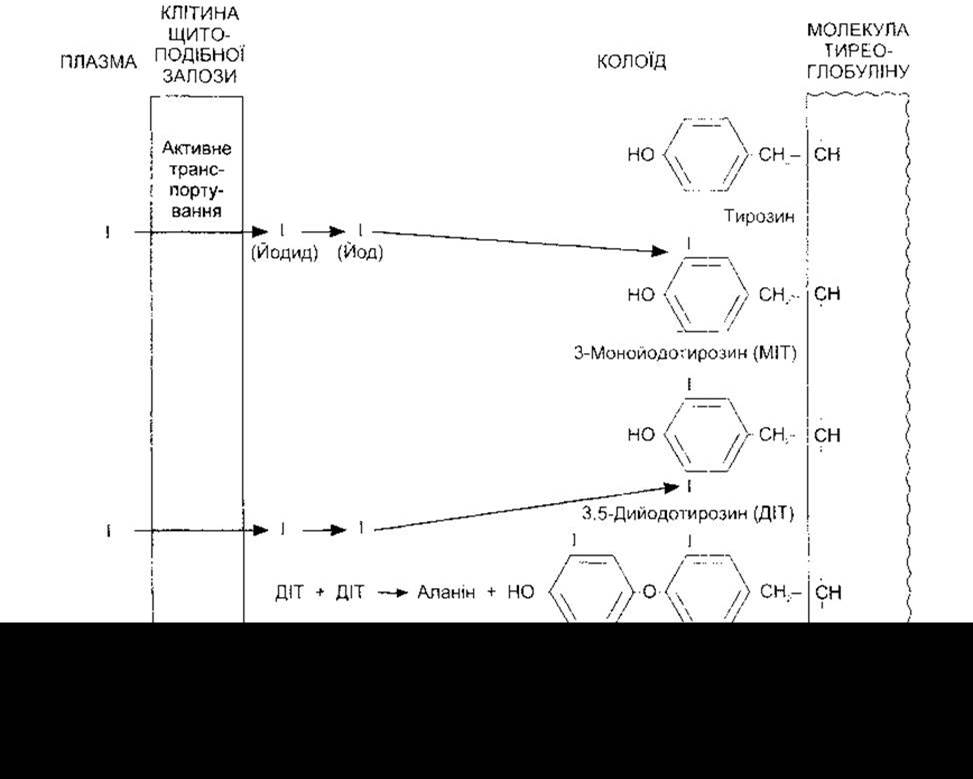
Рис. 18-6. Схема біосинтезу тиреоїдних гормонів. Йодування тирозину відбувається на апікальній поверхні клітин щитоподібної залози, тоді як молекули з’єднуються пептидними зв’язками з тиреоглобуліном.
Секреція
Щитоподібна залоза людини виділяє близько 80 мкг (103 нмоль) Т4, 4 мкг (7 нмоль) Т3 та 2 мкг (3,5 нмоль) ОТ3 за день (рис. 18-7), проте MIT та ДІТ не секретує. Клітини щитоподібної залози поглинають колоїд шляхом ендоцитозу (див. Розділ 1). Таке постійне поглинання на краю колоїду утворює лакуни реабсорбції, які видно в активних залозах (див. рис. 18-2). У клітинах глобули колоїду зливаються з лізосомами (див. рис. 18-3). Пептидні зв’язки між йодованими залишками та тиреоглобуліном руйновані протеазами в лізосомах, а Т4, Т3, ДІТ та MIT вивільняються в цитоплазму. Йодовані залишки тирозину дейодує мікросомальна йодотирозиндейодаза. Цей ензим не атакує йодовані тироніни, і Т4 та Т3 виходять у кров. Йод, вивільнений унаслідок дейодування MIT і ДІТ, залоза використовує повторно, що забезпечує вдвічі більше йодиду для синтезування гормонів, ніж дає йодидна помпа. У пацієнтів і суб’єктів, у яких спадково нема йодотирозиндейодази, MIT та ДІТ з’являються в сечі і простежуються симптоми нестачі йоду (див. нижче).
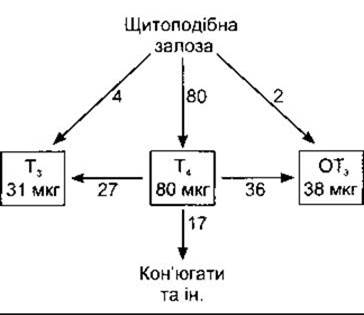
Рис. 18-7. Секреція та взаємоперетворення тиреоїдних гормонів у нормальних дорослих людей. Значення наведені у мікрограмах за день. Зауважте, що більшість Т3 та ОТ3 утворюється з Т4 шляхом дейодування в тканинах і лише невеликі кількості секретує щитоподібна залоза.