БІОТЕХНОЛОГІЯ - В. Г. Герасименко - 2006
Частина І. Загальна біотехнологія
Розділ 3. ОСНОВИ МОЛЕКУЛЯРНОЇ БІОЛОГІЇ
3.2. БІОСИНТЕЗ БІЛКА І ЙОГО РЕГУЛЯЦІЯ
3.2.1.Генетичний код
Про те, що ДНК є генетичним матеріалом, стало відомо завдяки дослідженням Ейвері О.Т. та співробітників (1944), що викликали трансформацію бактерій очищеними екстрактами ДНК пневмококів, а також роботами Херші А.Д. і Чейза М., які дослідили, що при зараженні бактеріальної клітини усередину проникає тільки ДНК бактеріофага, а його білкова оболонка залишається зовні. Зусиллями вчених (біохіміків, біофізиків, генетиків, хіміків і ін.) на початку 50-х років ХХ ст. була розшифрована просторова структура ДНК; хімічний склад цих макромолекул був визначений трохи раніше. Приблизно в цей же час вдалося визначити послідовність амінокислот у білку інсуліні, що складається тільки з 51 амінокислотного залишку. Таким чином, було переконливо доведено, що ДНК — це довгий нерозгалужений полімер, який складається з повторюваних у різній послідовності чотирьох мономерних структур — дезоксирибонуклеотидів, азотисті основи яких представлені аденіном (А), цитозином (С), гуаніном (G) і тиміном (Т). Мононуклеотиди з’єднані між собою ковалентними фосфодіефірними зв’язками, що йдуть від 5'-атома вуглецю одного залишку дезоксирибози до 3'-атому вуглецю наступного пентозного залишку і утворюють ланцюг — лінійну послідовність.
Методом рентгеноструктурного аналізу було встановлено, що ДНК має форму спіралі, яка складається з двох ланцюгів, розташованих таким чином, що азотисті основи виявляються усередині подвійної спіралі (сходинки кручених сходів), а де- зоксирибозофосфатний каркас виявляється зовні (поручні цієї драбини). Оптимальне упакування лінійних послідовностей мономерів у полінуклеотидній структурі подвійної спіралі досягається за рахунок взаємодії однієї великої пуринової основи (аденіну чи гуаніну), кожна з яких утворена шляхом конденсації шестичленного і п’ятичленного гетероциклів, з меншою за розміром піримідиновою основою (тиміном чи цитозином), які є шестичленними гетероциклами.
Результати модельних дослідів показали, що між гуаніном (G) і цитозином (С), а також між аденіном (А) і тиміном (Т) утворюється більше ефективних водневих зв’язків, чим за будь- яких інших сполучень нуклеотидів. Комплементарне спарювання А з Т і G із С в подвійній спіралі ДНК пояснило раніше отримані результати біохімічних досліджень ДНК про кількісну рівність А з Т і G із С, тобто співвідношення між азотистими основами в наведених парах у всіх досліджених ДНК складало 1:1.
У результаті біохімічного аналізу білків, що є продуктами мутантних генів, було показано, що послідовність чотирьох мономер- них структур (аденіну, гуаніну, тиміну і цитозину) у ДНК і двадцятьох амінокислотах у білках колінеарна, тобто послідовність нуклеотидів у ділянці ДНК, що кодує білок, відповідає послідовності амінокислот у цьому білку. Отже, такий стан справ порушив питання, яке стало головним у молекулярній біології, про механізм такого біохімічно складного перетворення, як переведення послідовності нуклеотидів ДНК у послідовності амінокислот білка. Потік інформації від ДНК до білка символічно можна подати так:
![]()
Подана схема привернула увагу вчених і надалі сприяла стрімкому розвитку біохімічної генетики. Синтез молекул РНК називається транскрипцією ДНК; утворена на матриці ДНК на одному з її ланцюгів, РНК-копія містить у собі увесь обсяг інформації цієї ділянки ДНК; РНК зберігає здатність до утворення водневих зв’язків між комплементарними основами, тому що урацил, присутній у РНК замість тиміну, спарюється з аденіном так само, як і тимін. Натомість транскрипція відрізняється від реплікації. РНК-копія після завершення її синтезу звільняється від ДНК-матриці, слідом за чим відбувається відновлення вихідної подвійної спіралі ДНК; знову синтезовані молекули РНК мають одноланцюгову структуру, вона коротше ДНК і відповідає довжині тієї ділянки ДНК, якої достатньо для кодування одного чи декількох білків. Одні ділянки ДНК (гени) використовуються для синтезу РНК тисячі разів, тоді як інші не транскрибуються зовсім.
У клітинах еукаріот багато з тих молекул РНК, що утворилися під час транскрипції, перш ніж перетворитися на інформаційну РНК (іРНК) і потрапити в цитоплазму, піддаються значним хімічним змінам. У свою чергу, у цитоплазмі на кожній молекулі іРНК можуть синтезуватися тисячі копій відповідного поліпептидного ланцюга. Якщо при цьому врахувати, що швидкість, з якою проходить процес білкового синтезу, надзвичайно висока (поліпептидний ланцюг, що складається зі ста амінокислотних залишків, у клітині кишкової палички створюється протягом 5 с), то інформація, що міститься у невеликій ділянці ДНК, здатна реалізуватися у вигляді великої кількості певного білка. Так, на матриці одного гена, що кодує фіброїн, відбувається синтез 104 молекул іРНК, на кожній з яких одержується 105 молекул фіброїну, що є основним компонентом шовку. У цілому за чотири доби одна клітина шовкосинтезуючої залози виробляє 109 молекул фіброїну (Албертс Б. і ін., 1986).
Правила переведення послідовності нуклеотидів, що входять у полінуклеотидну структуру ДНК, в амінокислотну послідовність білків (генетичний код) були розшифровані на початку 60-х років ХХ ст. Ніренбергом М., Маттеї Г., Ледером Ф. та іншими дослідниками. У більш ранніх генетичних експериментах було показане кодування амінокислот триплетами ну- клеотидів (кодонами). У цьому випадку з чотирьох азотистих основ (А, Т, G, С) можна скласти 64 (43) різні триплетні комбінації, яких цілком достатньо для кодування 20 амінокислот. Якщо ж з цього набору скласти сполучення по два нуклеотиди (дублетний код), то цієї кількості явно бракує для кодування всього набору амінокислот. Інкубацією у пробірках суміші, що складається з різних синтетичних полірибонуклеотидів, екстракту з кишкової палички, двадцяти амінокислот, з яких тільки одна мала радіоактивну мітку, вдалося установити весь набір триплетів для кодування усіх амінокислот. Крім того, за допомогою тринуклеотидів з відомою послідовністю азотистих основ була розшифрована нуклеотидна послідовність у всіх кодонах, що обумовлює зв’язування різних аміноацил-тРНК.
Варто також відзначити роботи Корани Х.Г., який запропонував метод хімічного синтезу полі- та олігонуклеотидів, і Холлі Р.У., що розшифрував структуру тРНК з антикодовою ділянкою.
Донедавна наявний фактичний матеріал свідчив про універсальність генетичного коду, тобто у всіх організмах — вірусах, прокаріотах і еукаріотах ті самі нуклеотидні триплети кодували однакові амінокислоти. В останні роки при вивченні процесу біосинтезу білка в мітохондріях були виявлені відхилення від універсального коду (табл. 3.3).
Таблиця 3.3.
Невідповідність генетичного коду мітохондрій універсальному коду (за Бохінські Р., 1987)
Кодони |
УГА |
АУА |
АГУ |
АГГ |
АУУ |
Універсальний код |
Термінація |
Іле |
Apr |
Apr |
Іле |
Мітохондріальний код |
Три |
Мет і ініціація |
Термінація |
Термінація |
Іле і, можливо, ініціація |
Для генетичного коду характерна ще його виродженість; у даному випадку це означає, що амінокислоті відповідає більш ніж один кодон. Наприклад, аргініну, лейцину і серину відповідає по шість; гліцину, проліну, валіну, тирозину й аланіну — по чотири, а триптофану і метіоніну — по одному кодону. Внаслідок виродженості генетичного коду помилки, що виникають при реплікації і транскрипції в деяких випадках не супроводжуються перекручуванням генетичної інформації і порушенням експресії, що має біологічне значення. В усіх випадках дво-, три- і чотириразової виродженості зміна відбувається тільки в третьому нуклеотиді триплету. Так, якби аланін кодувався тільки одним триплетом ГЦУ, то будь-яка зміна нуклеотидної послідовності в ньому при реплікації або транскрипції неминуче супроводжувалася б заміною у відповідній поліпептидній структурі аланіну іншою амінокислотою з усіма наслідками, що з цього випливають. Однак завдяки чотириразовій виродженості тільки заміни, що стосуються перших двох нуклеотидів кодона, ведуть до зміни його значення. Специфічність кодонів визначається головним чином його першими двома нуклеотидами. Що стосується третього нуклеотида, який займає положення на 3'-кінці олігонуклеотидної структури, то його специфічність виражена слабше (Ленінджер А., 1985; Бохінські Р, 1987).
Точність синтезу поліпептидного ланцюга досягається за рахунок комплементарного розпізнавання азотистих основ 5/^3/-орієнтованої послідовності кодона іРНК, що має 3'→5'- напрямок послідовності азотистих основ антикодону тРНК.
Слід зазначити, що кількість каталізуючих реакцію активування амінокислот аміноацил-тРНК-синтетаз відповідає кількості різних видів амінокислот, з яких синтезуються білки; що стосується тРНК, то їхня кількість як мінімум повинна досягати 32, тому що деякі амінокислоти здатні взаємодіяти з двома, а то й з трьома різними тРНК, які, у свою чергу, пізнають і зв’язують один, два чи навіть три кодони іРНК. Кодон-антикодонове пізнавання (кодон розташовується на іРНК і має 5'→3'-орієнтацію, антикодон — на тРНК і орієнтований у напрямку 3'→5') передбачає відхилення від класичної взаємодії й утворення водневих зв’язків між парами азотистих основ А — Т, G — С і А — U у ДНК і РНК. Це відхилення сформульоване Кріком Ф. у гіпотезі коливань, біологічний зміст якої має багато спільного з явищем виродженості генетичного коду. Гіпотеза коливань зводиться до здатності третьої азотистої основи, розташованої з боку 5'-кінця антикодону (тРНК), змінювати своє просторове положення, натомість як дві перші основи, що знаходяться в 3'-кінці антикодону, фіксовані більш жорстко. Зміна просторової орієнтації, що знаходиться на 5'-кінці антикодону азотистої основи обумовлює його здатність до утворення не відповідних класичним (А — Т, G — С і А — U) взаємодій між парами основ. Установлено, що третім від 3'-кінця антикодону може бути U, G чи І (рибонуклеозид інозин, у якому азотистою основою виступає гіпоксантин, що утворюється при відщепленні від аденіну його 6-аміногрупи). В табл. 3.4 подано можливі варіанти поєднання пар основ у зв’язку з явищем коливань.
Таблиця 3.4.
Можливі поєднання пар основ між 5'-кінцем антикодону (тРНК) і 3'-кінцем кодону (іРНК), визначені гіпотезою коливань
(за Бахінські Р., 1987)
Основа на 5'-кінці антикодону (тРНК) |
Основа 3'-кінці кодону (іРНК) |
I G U A* C* |
A, C або U C або U A або G U G |
* Гіпотеза не передбачає нових комбінацій, якщо ці основи знаходяться в антикодоні
Як видно з табл. 3.4, якщо в антикодоні в положенні коливання знаходиться гіпоксантин (у складі інозину), можливе розпізнавання й утворення водневих зв’язків трьох пар: І — А, І — С і І — U (але така комплементарна взаємодія виявляється більш слабкою порівняно з виникаючою взаємодією при утворенні звичайних пар G — С i А — U); у випадку перебування у позиції коливання G чи U кількість можливих сполучень обмежується двома: G — С, G — U і U — А, U — G. Нові комбінації пар основ не виникають, якщо такими, що коливаються, є аденін і цитозин. У цьому випадку відбувається утворення зв’язків за класичним принципом: А — U; С — G (рис. 3.7).
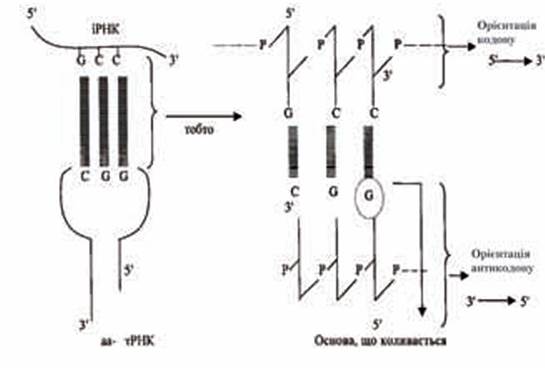
Рис. 3.7. Схема кодон-антикодонових взаємодій у світі гіпотези коливань
(за Бохінські Р. 1987)
Утворення слабких водневих зв'язків при колон-антикадоновому дізнаванні можна показати на прикладі однієї з аргінінових тРНК, антикодон якої (5') І — С — G (3') здатний взаємодіяти з трьома різними аргініновими кодонами:
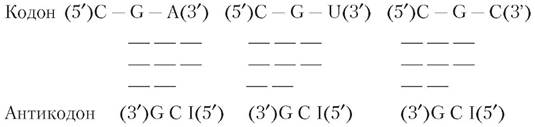
Дві перших основи кодонів (С — G) утворюють міцні (позначені трьома рисками) уотсон-кріковські пари з відповідними азотистими основами антикодону. Азотисті основи, що знаходяться у третьому положенні, (А, U, С) аргінінових кодонів утворюють слабкі водневі зв’язки (дві риски) із залишком інозину (І) в антикодоні. На підставі цього й інших прикладів кодон-антикодонових взаємодій Крік Ф. дійшов висновку, що основи більшості кодонів, які знаходяться у третьому положенні, мають деякий ступінь свободи при утворенні пари з відповідною азотистою основою антикодонів, тобто, за термінологією Кріка Ф., основи, що коливаються. Біологічний зміст явища полягає в тому, що воно дозволяє звести до мінімуму виникаючі помилки. Завдяки неміцності зв’язку, що утвориться між основою, яка коливається, відповідною основою антикодону, тРНК легше звільняється з комплексу іРНК у процесі білкового синтезу. У випадку залучення в сильну уотсон-кріковську кодон-антикодонову взаємодію усіх трьох пар основ міцність зв’язку стала б моментом, що лімітує швидкість білкового синтезу через уповільнення процесу вивільнення тРНК із комплексу з іРНК (Ленинджер А., 1985; Бохінські Р., 1987).