БІОТЕХНОЛОГІЯ - Іншина Н.М. - 2009
РОЗДІЛ 1. ГЕННА ІНЖЕНЕРІЯ
Бактерії, що стимулюють ріст рослин
Більшість грунтових мікроорганізмів стимулюють ріст рослин за такими механізмами:
- фіксація атмосферного азоту;
- утворення легкозасвоюваних хелатованих форм Форуму і Фосфору, їх поглинання з ґрунту і транспортування у рослини;
- синтез речовин, що пригнічують ріст фітопатогенів;
- синтез фітогормонів;
- синтез мікробних інсектицидів;
Бактерії, що стимулюють ріст рослин, можуть використовувати один або кілька таких механізмів.
Молекулярні механізми фіксації азоту
Цінною властивістю бактерій є диазотрофність - здатність до фіксації атмосферного азоту. За допомогою азотфіксуючих бактерій щорічно 17,5·107 т молекулярного азоту атмосфери перетворюється в органічні сполуки. Усі мікроорганізми, що здійснюють азотфіксацію, є прокаріотами. Жоден еукаріот не здатний зв’язувати атмосферний азот. До азотфіксуючих бактерій належать: Rhizobium, Brady rhizobium, Frankia, Azospirillum, Azotobacter і ціанобактерії. З допомогою методів генної інженерії створено бульбочкові бактерії з підвищеною здатністю до фіксації атмосферного азоту.
Фіксацію атмосферного азоту у бактерій здійснює фермент нітрогеназа. Молекулярні основи фіксації азоту були досліджені на прикладі бактерії Klebsiella pneumoniae, що мешкає у грунті, воді, кишечнику людини. Klebsiella pneumoniae може слугувати модельною системою для вивчення нітрогенази бактерій Rhizobium і Bradyrhizobium.
Нітрогеназна азотфіксуюча система є складним мультиферментним комплексом. Усі відомі нітрогенази містять два компоненти. Компонент І - це комплекс із двох α-субодиниць (50 кДа) і двох β-субодиниць (60 кДа), 24 молекул Феруму, 2 молекул Молібдену і залізомолібденового кофактора (FeMoCo). Компонент II складається із двох α-субодиниць (32 кДа) і невідомого числа молекул Феруму. Для фіксації азоту необхідні обидва компоненти нітрогенази, комплекс йонів Mg2+ й АТФ та джерело відновних еквівалентів. Схематично процес фіксації азоту можна описати таким рівнянням:
N2 + 8Н+ + 8 е + 16 MgATФ = 2 NH3 + Н2 + 16 MgAТФ + 16 Н3РО4.
Гени азотфіксації позначають як nif-гени, вони кодують приблизно 20 різних білків (табл. 1.5). тому створення з допомогою методів генної інженерії рослин, здатних самостійно фіксувати азот, є практично неможливим.
Бульбочкові бактерії в симбіозі з бобовими рослинами відновлюють азот до аміаку, який може використовуватися для синтезу глутаміну, глутамінової кислоти і білка.
Нітрогеназа каталізує утворення газоподібного водню. Деякі штами Rhizobium синтезують фермент гідрогеназу, який каталізує перетворення in vivo Н2 у Н+, що збільшує ефективність фіксації азоту. За допомогою генної інженерії можна підвищити азотфіксуючу здатність бактерій Rhizobium шляхом уведення генів ферментів гідрогеназ. Гідрогеназна система може застосовуватися для таких процесів:
- підвищення ефективності фіксації азоту;
- перетворення і запасання сонячної енергії;
- регенерації кофакторів у промислових ферментативних процесах;
- синтезу специфічних хімічних сполук;
реакторів атомних електростанцій.
Таблиця 1.5
Гени Klebsiella pneumoniae, що беруть участь у фіксації азоту
nif-Ген |
Білок (функція) |
D |
α-Субодиниця компонента І нітрогенази |
К |
β-Субодиниця компонента І нітрогенази |
Η |
Компонент II нітрогенази |
F |
Флаводоксин |
J |
Піруват: флаводоксин оксидоредуктаза |
O, B, N, E, V |
Синтез FeMoCo |
M |
Процесинг редукгази нітрогенази |
A |
Активатор |
L |
Репресор |
S |
Процесинг компонента І |
W, Z, T, Y, U, X |
Невизначені функції |
Бактеріальні добрива на основі бульбочкових бактерій
Бактеріальні добрива збагачують ризосферу рослин корисними мікроорганізмами, що забезпечує покращання структури ґрунту, накопичення у ньому поживних речовин, мінералізацію органічних сполук і, як наслідок, підвищення родючості фунту і врожайності рослин. Мікроорганізми бактеріальних добрив забезпечують рослини не тільки мінеральними речовинами, а й фізіологічно активними сполуками: фітогормонами, вітамінами.
У сільському господарстві найчастіше застосовують бактерії двох родів: Rhizobium і Bradyrhizobium. У результаті симбіозу з бобовими рослинами бактерії одержують органічні речовини, а рослини - NH4+. Кожен вид бульбочкових бактерій специфічний відносно невеликої кількості видів рослин.
Уперше культура бульбочкових бактерій була одержана у 1911 р. на бактеріально-агрономічній станції у Москві. На основі бульбочкових бактерій розроблено такі бактеріальні добрива: нітрагін, ризоторфін, азобактерин, фосфобактерин, екстрасол. В 1 г нітрагіну міститься приблизно 9 млрд бактерій. Обробка насіння рослин нітрагіном підвищує врожайність на 15-25%. Для виробництва азобактєрину використовують бактерії Azotobacter chroococcum. Azotobacter фіксує атмосферний азот, продукує вітаміни (В5, В3, В6, біотин), стимулятори росту рослин (гетероауксин, гіберелін), фунгіцидні речовини, що пригнічують ріст небажаних мікроскопічних грибів. Фосфобактерин створений на основі бактерій Bacillus megaterium var.phosphaticum, які перетворюють складні фосфорорганічні сполуки (нуклеїнові кислоти, нуклеопротеїди) і важкозасвоювані мінеральні фосфати в доступну для рослин форму. 1 г препарату фосфобактерину містить близько 8 млрд бактерій. Мікроорганізмами обробляють насіння і рослини перед висаджуванням у ґрунт.
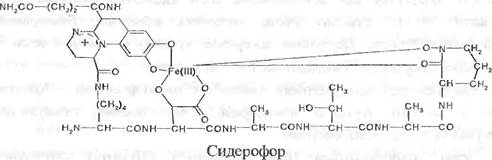
Мікробіологічний синтез сидерофорів
Сидерофори — де речовини, що хелатують залізо. Як відомо, Ферум присутній у грунті переважно у складі сполук Fe3+, що нерозчинні у воді. Грунтові мікроорганізми синтезують і декретують залізозв’язуючі сполуки з молекулярною масою 400 - 1000 Да.
Сидерофори ефективно зв’язують Fe3+ і забезпечують надходження Форуму до клітин мікроорганізмів, які, у свою чергу, постачають його у рослини.
ВільноживучІ бактерії Azospirillum здатні стимулювати ріст рослин за рахунок синтезу і секреції органічних кислот, які розчиняють і зв’язують мінеральні речовини.
Синтез речовин, що пригнічують діез фітопатогенів
Фітопатогени можуть зменшити врожай сільськогосподарських культур на 25 - 100%. Деякі мікроорганізми синтезують сполуки, що пригнічують ріст фітопатогенів. Так, наприклад, бактерії роду Pseudomonas синтезують антибіотики: агроцин 84, агроцин 434, гербіколін, феназини, піролнітрил, піолутеорин.
Деякі бактерії мають протигрибкову дію, оскільки синтезують ферменти (хітиназа, Р-1,3-глюканаза), що руйнують клітинну стінку грибів. В одному експерименті вдалося знизити частоту захворювання рослин фітопатогенними грибами Rhizoctonia solani, Sclerotium rolfsii, Pythium ultium за допомогою штаму Pseudomonas cepacia, що синтезує фермент р-1,3-глюканазу.
Ген хїтинази, виділений з бактерії Serrat ia marcescens, був перенесений у штам Pseudomonas fluorescwcns. Одержаний штам мікроорганізмів секретував хітиназу й ефективно пригнічував розмноження фітопатогенного гриба Rhizoctonia solani.
Синтез фітогормонів
Бактерії можуть стимулювати ріст рослин через зміну їх гормонального балансу. Бактерії синтезують фітогормони, що стимулюють ріст рослин, прискорюють поділ їх клітин і диференціювання. Основними представниками фітогормонів є ауксини і цитокініни.
Назва «ауксини» об’єднує ряд речовин — регуляторів росту. Найважливішими представниками ауксинів є індолілоцтова, ікдолілпропіонова, 2,4-дихлорфеноксиоцтова кислоти.
Ауксини були відкриті на початку Другої світової війни. Спочатку ауксини отримували із верхівки колеоптилю - чохла, що захищає перший молодий листок. За 10 днів 8 лаборанток німецького біохіміка Кегля переробили 100 тисяч проростків кукурудзи й одержали ауксин у кількості, достатній для встановлення його кислотної природи. Щоб одержати 250 мг ауксину таким способом, необхідно безперервно працювати 400 років. Доступним джерелом ауксину є людська сеча. За добу з організму людини виводиться 1 - 2 мг ауксину.
Ефективними продуцентами ауксинів є мікроорганізми - дріжджі, гриби і бактерії. Ауксини прискорюють ріст рослин, стимулюють формування їх кореневої системи.
Деякі мікроорганізми (гриб Gibborolla fujukuroi) синтезують терпеноїди - гібереліни. Гібереліни стимулюють не лише ріст, а й цвітіння рослин. їх застосовують для прискорення проростання ячменю під час виготовлення солоду, для підвищення врожайності винограду. Пізніше були відкриті сполуки, що стимулюють поділ клітин, - цитокініни. Цитокініни є похідними 6-амінопурину. У присутності ауксинів вони індукують поділ клітин, стимулюють процеси синтезу РНК і білків.
Фітогормони характеризуються високою ефективністю дії, вони справляють свій вплив у дуже низьких концентраціях. Наприклад, 1 г гетероауксину достатньо для стимулювання росту 1013 рослин. Якщо висадити таку кількість рослин у ґрунт (із розрахунку 1 рослина на 1см2), то площа поля становитиме 900 км2. Для приготування розчину, що містить 1 г гетероауксину, необхідно використати 200 млрд л води.
Синтез мікробних інсектицидів
Найвідоміший хімічний інсектицид ДДТ (дихлордифенілтрихлоретан) був синтезований у 70-х роках XIX ст. З 30-х років XX ст. ДДТ застосовують для боротьби з комахами-шкідниками. Як і більшість хлорорганічних сполук, ДДТ має паралізуючу дію на нервову систему і м’язову тканину комах. Синтезовано і широко використовують й інші хлорорганічні сполуки: дильдрин, альдрин, хлордан, ліндан, токсофен. Інший клас хімічних інсектицидів — фосфорорганічні сполуки (малатион, паратион, диазинон). Вони інгібують фермент ацетилхолінестеразу, що гідролізуе нейромедіатор ацетилхолін. Фосфорорганічні інсектициди порушують функціонування нейронів у комах.
Хімічні інсектициди шкідливо впливають на людину і тварин. Хлорорганічні сполуки, зокрема ДДТ, зберігаються в екосистемах тривалий час — від 15 до 20 років. З часом комахи-шкідники виробляють стійкість до багатьох хімічних інсектицидів, тому ці сполуки стали використовувати у вищих концентраціях. Хімічні інсектициди не мають вибіркової дії, тому разом із шкідниками знищують корисних комах. Хімічні інсектициди акумулюються в жировій тканині багатьох організмів. Так, унаслідок отруєння хімічними інсектицидами в Північній Америці були винищені такі види птахів: сапсани, бурі пелікани, білоголові орлани. З урахуванням цього здійснюється інтенсивний пошук альтернативних способів контролю чисельності комах-шкідників.
Мікробні інсектициди, на відміну від хімічних, не справляють шкідливого впливу на навколишнє середовище. Інсектициди, що синтезують мікроорганізми, високоспецифічні, діють лише на певні види шкідливих комах. Мікробні інсектициди підлягають швидкій біодеградації, у комах не виробляється стійкість до біоінсектииидів. Проте широке використання цих сполук обмежується високою вартістю їх одержання. Цю проблему можна подолати за допомогою генної інженерії. Для одержання мікробних інсектицидів використовують віруси, гриби, найпростіші, бактерії.
У вітчизняному біотехнологічному виробництві виділяють 3 групи інсектицидних препаратів:
1) бактеріальні препарати на основі Bacillus thuringiensis (ентобактерин-3, екзотоксин, інсектин, токсобактерин);
2) грибні препарати (боверин);
3) препарати на основі вірусів ядерного поліедра (вірин).
Бактеріальні ентомопатогенні препарати застосовуються найчастіше. Вони характеризуються такими особливостями дії:
- висока вірулентність щодо комах-шкідників;
- безпечність для довкілля;
- висока швидкість дії на шкідників.
Найбільш вивчені ентомопатогенні бактерії Bacillus thuringiensis. Потрапляючи в організм комах, вони продукують токсини:
- α-екзотоксин (або фосфоліпаза С) індукує розпад фосфоліпідів у тканинах комах;
- β-екзотоксин інгібує процес транскрипції (синтез РНК);
- γ-екзотоксин - малодосліджений фермент;
- δ-ендотоксин - 8-гранний кристал, який під час потрапляння у кишечник комах розпадається на молекули протоксина, що пошкоджує середній відділ кишечнику.
ГІротоксин Bacillus thuringiensis спричиняє загибель комахи вже через 15 хв дії. Токсини Bacillus thuringiensis швидко руйнуються. Бактерії Bacillus thuringiensis антагоністичні до 130 видів комах.
Бактеріальні ентомопатогенні препарати застосовують шляхом розпилювання водної емульсії (на 1 га овочевих культур використовують 1 - 3 кг препарату, для садових культур - 3 - 5 кг). 1 т мікробного препарату достатньо, щоб знищити шкідників на 300 га лісу, бурякових чи бавовникових полів.
Клоновані гени різних токсинів Bacillus thuringiensis, які локалізовані у плазмідах. Один з таких генів був уведений у фотосинтезугочі ціанобактерії Synechocystis i Synechococcus. Цей підхід виявився ефективним для доставки токсинів Bacillus thuringiensis в організм комахи- мішені. Однак інсектициди Bacillus thuringiensis під час нанесення на листки і стебла не діють на комах, що пошкоджують корені рослин. Ген токсину В. thuringiensis увели в штам одного з видів бактерій, що мешкають у ризосфері — шарі грунту поблизу коренів. Такі рекомбінантні бактерії секретують інсектицид прямо в ризосферу і захищають корені рослин від комах-шкідників. Також ген токсину В. thuringiensis був вбудований у хромосомну ДНК штаму Pseudomonas fluorescens. що утворює колонії на коренях кукурудзи.
Грибні ентомопатогенні препарати спричиняють у комах мікози. Особливості дії мікроскопічних грибів:
- ураження комах відбувається через кутикулу;
- комахи інфікуються у фазі розвитку лялечки та імаго;
- висока швидкість росту ї репродуктивна здатність грибів (у вигляді спор можуть довго зберігатися, не втрачаючи ентомопагогенної дії);
- висока специфічність - вірулентність залежить від штаму гриба.
Дія грибного препарату на комаху розпочинається після потрапляння спор у порожнину її тіла через шкірні покриви. В організмі комах спора проростає в гіфу, потім розростається міцелій, який заповнює все тіло комах, продукуються токсини. Ріст гриба триває доти, поїси всі тканини комахи, насамперед м’язова, не будуть зруйновані.
У промисловому виробництві грибних інсектицидів найчастіше використовуються штами трьох родів: Beaveria. Metarrhizium, Entomophtora. На основі гриба Beauveria bassianaбув створений препарат боверин (1 г препарату містить 1,5-6 млрд конідіоспор).
Вірусні ентомопатогенні препарати володіють найбільшою специфічністю відносно комах, тому вони безпечні для людини, флори і фауни. Ці препарати діють лише на один вид комах. Віруси стійкі до несприятливих умов середовища, вони можуть зберігати життєздатність протягом багатьох років. Інфікування вірусами відбувається під час живлення комах. Віріони через стінку кишечнику проникають у клітини, де здійснюється їх реплікація.
Для боротьби з комахами- шкідниками використовують бакуловіруси - паличкоподібні віруси з дволанцюговим ДНК-геномом. Стійкість до бакуловірусів розвивається дуже рідко, оскільки бакуловіруси протягом тисячоліть еволюціонували разом зі своїми комахами-хазяїнами. Унаслідок дії токсинів бакуловірусів комахи можуть загинути у період від кількох днів до тижнів. Щоб прискорити цей процес, були зроблені спроби підвищити вірулентність бакуловірусів шляхом уведення чужорідних генів, експресія яких призводить до загибелі комах. В один із штамів бакуловірусу був уведений ген нейротоксину скорпіона Androctonus australis Hector. Цей токсин блокує транспорт іонів Na+ у нейронах комах, що призводить до паралічу і смерті. Уведення гена вдвічі підвищило вірулентність бакуловірусів. Рєкомбінантний бакуловірус не тільки прискорював загибель комах, а й знижував їх здатність пошкоджувати рослини. Основною перешкодою для широкомасштабного застосування бакуловірусів є висока вартість їх препаратів.
Ураховуючи шкідливий вплив хімічних інсектицидів на довкілля, мікробіологічні інсектициди є найперспективнішими для застосування у сільському господарстві.