Молекулярная биология клетки - Том 2 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1993
Рост и деление клеток
Дрожжи как модельная система
Дрожжи являются одноклеточными грибами и составляют большую группу довольно разнородных организмов. Поскольку они размножаются почти так же быстро, как бактерии, и размеры их генома меньше 1/1000 генома млекопитающих, они оказались чрезвычайно полезными для генетического анализа клеточной биологии эукариот. Хотя яйца Xenopus-исключительно ценный объект для изучения биохимических и цитофизиологических аспектов регуляции клеточного цикла, для генетических исследований этот объект неудобен. Напротив, работа с дрожжами открывает широкие возможности для идентификации, клонирования и описания генов, участвующих в контроле клеточного цикла. Мы будем здесь говорить о двух видах: о почкующихся дрожжах Saccharomyces cerevisiae, используемых пекарями и пивоварами, и делящихся дрожжах Schizosaccharomyces pombe. Клетки этих последних делятся симметрично на две одинаковые дочерние клетки, а у почкующихся дрожжей они делятся менее распространенным несимметричным образом: материнская клетка производит маленькую почку, которая растет и проходит оставшиеся фазы цикла, прежде чем окончательно отделиться от родительской клетки (рис. 13-16).

Рис. 13-16. Сравнение клеточных циклов делящихся и почкующихся дрожжей. У делящихся дрожжей (вверху) типичный цикл эукариотической клетки с фазами G1, S, G2 и М. Ядерная оболочка, однако, не разрушается: микротрубочки митотического веретена образуются внутри ядра и прикреплены к полюсным тельцам веретена на его периферии. Клетка делится надвое путем образования перегородки (называемой клеточной пластинкой). У почкующихся дрожжей цикл включает нормальные фазы G1 и S, однако состоящее из микротрубочек веретено начинает формироваться очень рано, во время фазы S, и поэтому нормальная фаза G2 отсутствует. В отличие от цикла делящихся дрожжей здесь во время митоза не происходит видимой конденсации хромосом и клетка делится путем почкования. Как и у делящихся дрожжей (но в отличие от клеток высших эукариот), ядерная оболочка во время митоза сохраняется.
Как полагают, эволюционные ветви, приведшие к почкующимся и к делящимся дрожжам, дивергировали сотни миллионов лет назад. Тем не менее жизненные циклы у тех и других сходны. Обе формы могут размножаться либо в диплоидном, либо в гаплоидном состоянии. Диплоидные клетки помимо деления обычным путем способны проходить через мейоз, образуя гаплоидные клетки (см. гл. 15); а гаплоидные клетки наряду с обычным делением могут попарно сливаться между собой с образованием диплоидных клеток (рис. 13-17; см. также разд. 10.3.2). Наличие гаплоидной фазы облегчает генетический анализ и позволяет выделять мутантов с утраченными функциями; у диплоидного организма такие мутации находились бы в рецессивном состоянии (как это имеет место у культивируемых клеток млекопитающих) и поэтому их было бы труднее выявлять и учитывать. У обоих видов дрожжей в контроле клеточного цикла важную роль играют питание и половой процесс; в связи с этим дрожжи послужат хорошим объектом для рассмотрения общего вопроса о том, как цикл деления регулируется факторами внеклеточной среды.
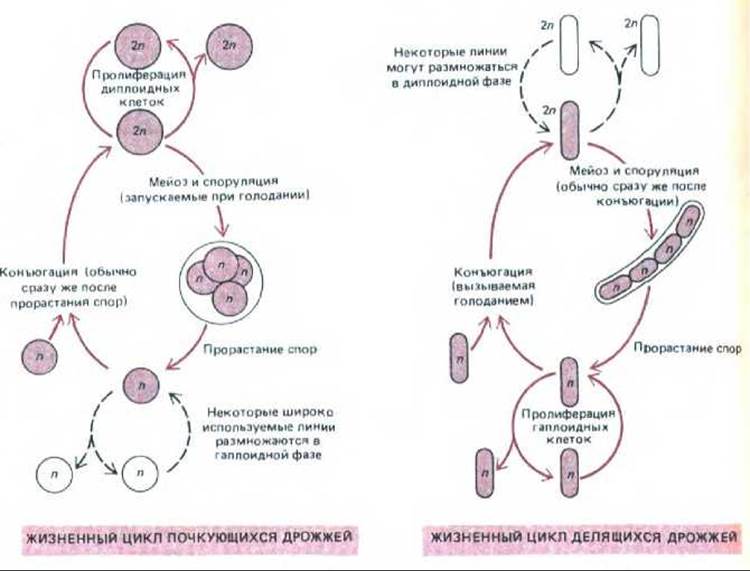
Рис. 13-17. Жизненные циклы почкующихся дрожжей (Saccharomyces cerevisiae) и делящихся дрожжей (Schizosaccharomyces pombe). Доли жизненного цикла, проводимые в гаплоидной и в диплоидной фазе, меняются от вида к виду и в зависимости от условий среды. При обилии пищи нормальные разновидности дикого типа почкующихся дрожжей размножаются как диплоидные клетки с продолжительностью жизненного цикла около двух часов. При голодании же они претерпевают мейоз с образованием гаплоидных спор, которые в благоприятных условиях прорастают, превращаясь в гаплоидные клетки. Эти клетки в зависимости от условий среды и генотипа либо делятся, либо сливаются (конъюгируют) в фазе G1, вновь образуя диплоидные клетки. Наоборот, делящиеся дрожжи обычно размножаются в гаплоидном состоянии; при недостатке пищи гаплоидные клетки сливаются с образованием диплоидных клеток, которые быстро проходят мейоз и споруляцию с восстановлением гаплоидной фазы. Наиболее широко используемые лабораторные штаммы почкующихся дрожжей - это мутанты, которые, подобно делящимся дрожжам, размножаются главным образом в гаплоидной фазе.
13-10
13.2.1. Каждая мутация, затрагивающая цикл деления дрожжевой клетки, останавливает или нарушает ход этого цикла в определенной его фазе [12, 13]
Чтобы идентифицировать гены, участвующие в контроле клеточного цикла, нужны соответствующие мутанты и способы получения от них потомства. Однако клетки с нарушенным механизмом клеточного цикла не способны размножаться. Выход из положения состоит в поиске условных мутантов, у которых дефект проявляется в фенотипе только при определенных условиях. Обычно ищут генный продукт, молекулярная структура которого слегка изменена так, что он утрачивает свою функцию в одном (рестриктивном) диапазоне температур, но сохраняет ее в другом (пермиссивном) диапазоне. Для таких термочувствительным мутаций низкие температуры обычно бывают пермиссивными, а высокие-рестриктивными. Таким образом, можно получить мутанта при низкой температуре, а затем, подняв ее, выключить измененный ген и исследовать мутантный фенотип.
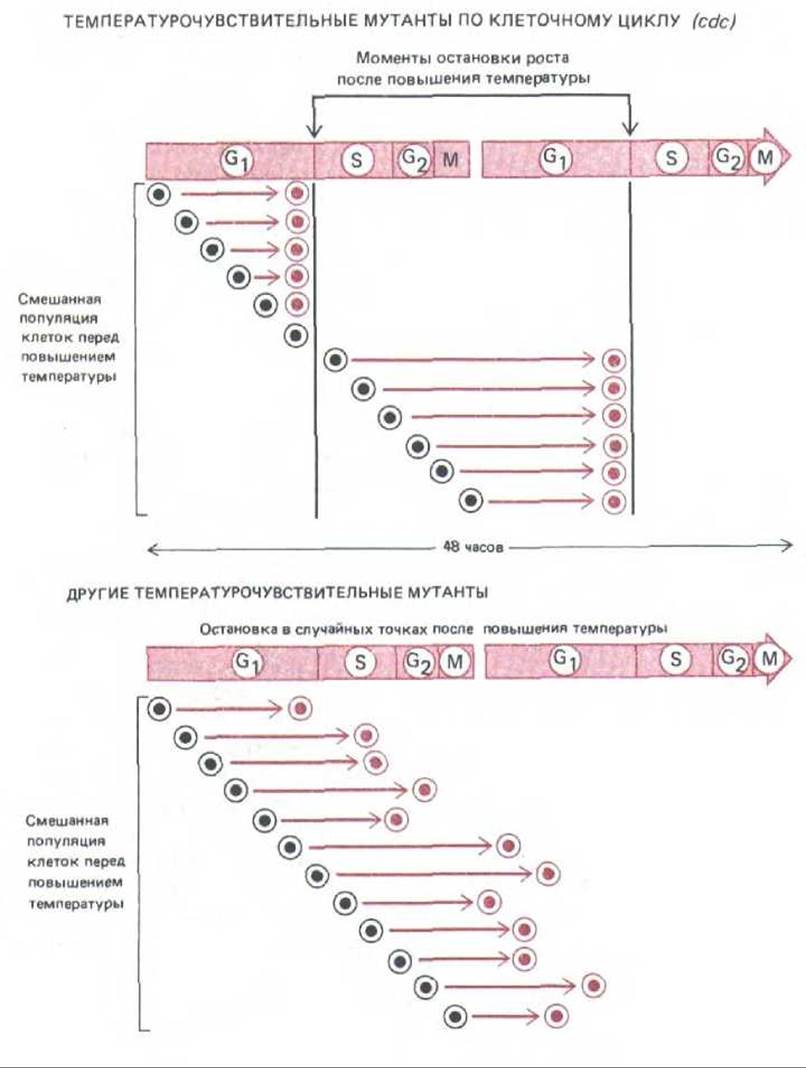
Рис. 13-18. Эти схемы показывают, как можно отличить термочувствительвого мутанта с измененным механизмом клеточного цикла (cdc) от других термочувствительных мутантов. При повышении температуры до рестриктивного уровня, когда продукт мутантного гена не может функционировать нормально, мутант будет продолжать свой клеточный цикл до тех пор, пока не дойдет до этапа, который он не в состоянии пройти (в данном случае это инициация фазы S). Поскольку, несмотря на блокаду цикла, клетка продолжает расти, мутанты cdc становятся ненормально большими (на схеме не показано). Между тем при других мутациях, вызывающих нарушение процессов, необходимых для роста на протяжении всего цикла (таких, как синтез АТР), клетка будет останавливаться в любой стадии цикла, как только она израсходует свои биохимические резервы.
Мутации, специфически влияющие на отдельные компоненты механизма клеточного цикла, нельзя обнаружить по одной лишь утрате способности мутантных клеток к делению, так как к этому будет приводить любой летальный дефект. Мутации цикла клеточного деления (cdc-cell-division-cycle) более достоверно выявляются по тому, как они блокируют или нарушают специфическую фазу клеточного цикла при пермиссивной температуре (рис. 13-18). У почкующихся дрожжей наличие и размеры почки служат простым визуальным индикатором, показывающим, какой этап клеточного цикла блокирован у данного мутанта cdc; в случае делящихся дрожжей нужны более сложные подходы с использованием методов анализа клеточного цикла, о которых говорилось выше.
У каждого из двух упомянутых видов дрожжей было идентифицировано от 40 до 50 генов cdc. В ряде случаев биохимический анализ позволил точно определить функцию генного продукта. Например, некоторые мутанты cdc, у которых цикл блокирован в S-фазе, оказались дефектными по генам, кодирующим ДНК-лигазу или ферменты, необходимые для синтеза предшественников ДНК. Как будет описано ниже, общим подходом для характеристики всех белков, кодируемых генами cdc, может служить метод рекомбинантной ДНК. Однако некоторые важные моменты можно выяснить даже без этой информации.
13-11
13.2.2. Дрожжевые мутанты cdc могут быть использованы для анализа сопряжения между событиями клеточного цикла
Если повысить температуру до рестриктивного уровня, у большинства мутантов cdc клеточный цикл останавливается на той стадии, на которой действует продукт гена cdc. Как правило, клетка теряет способность переходить к следующей стадии цикла, и это означает, что начало каждого процесса находится в зависимости от завершения предыдущего процесса. Таким образом, у дрожжей, как и у млекопитающих, большинство этапов клеточного цикла, по-видимому, связаны между собой как звенья единой цепи. Эта связь была более тщательно проанализирована в экспериментах с клетками, содержащими разные комбинации различных мутаций cdc. Как показали результаты, события хромосомного цикла образуют ряд зависимых друг от друга этапов, который не связан жестко с событиями цитоплазматического цикла (рис. 1319). Например, хотя цитокинеза не произойдет, если предотвратить деление ядра, тем не менее мутанты cdc, не способные пройти цитокинез из-за дефектов в механизме формирования почки, все же осуществляют повторные циклы синтеза ДНК и деления ядра. По-видимому, общим правилом не только для дрожжей, но и для клеток млекопитающих, насекомых и многих других организмов является то, что хромосомный цикл может продолжаться, даже если цитокинез предотвращен. В самом деле, и в ходе нормального развития таким способом часто возникают многоядерные клетки (разд. 16.5.2).
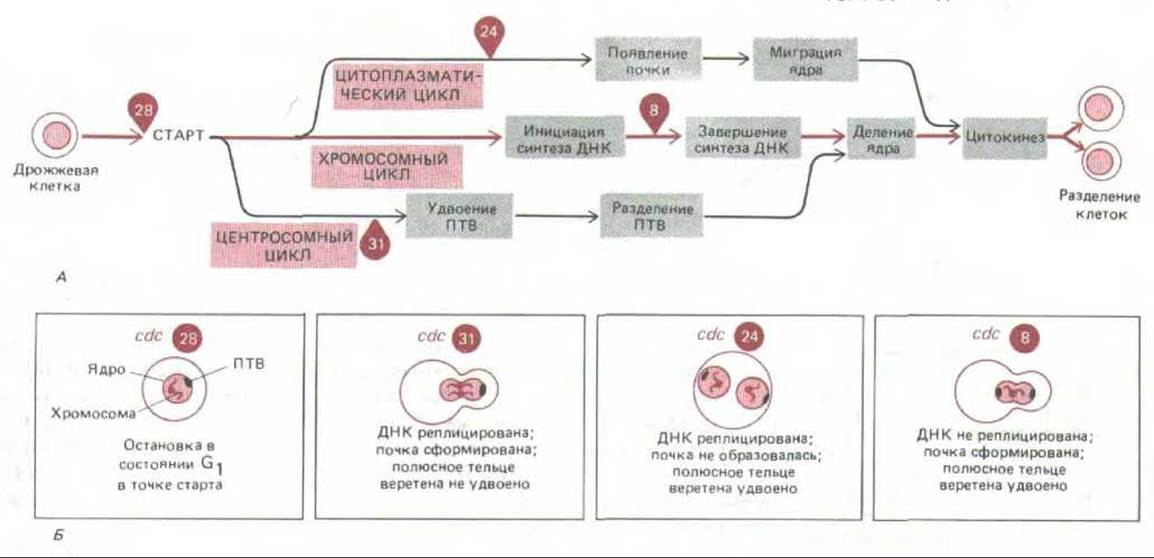
Рис. 13-19. Причинные связи между некоторыми событиями клеточного цикла и их отношение к генам cdc у почкующихся дрожжей. Полюсное тельце веретена (ПТВ) у дрожжей является эквивалентом центросомы. «Старт» означает точку бесповоротного вступления клетки в цикл деления и потерю возможности половой конъюгации (конъюгация может осуществляться только в фазе G1). A. Общая схема цикла; стрелка, идущая от события а (или от событий а к б) к событию в, означает, что событие е не может произойти раньше события а (или а и б вместе). Цифрами обозначены специфические мутации cdc, которые при рестриктивной температуре приводят к остановке клеток в данной точке цикла. Например, клетки с мутацией cdc8 останавливаются во время синтеза ДНК. Обратите внимание на то, что хромосомный цикл (цикл ДНК), цитоплазматический цикл (цикл формирования почки) и центросомный цикл (цикл ПТВ) частично независимы. Б. Блокированные состояния четырех мутантов cdc, указанных на схеме А, при высокой температуре. [По данным L. Н. Hartwell, J. Cell Biol. 77: 627-637, 1978; J. R. Prmgle, L.H. Hartwell. In: The Molecular Biology of the Yeast Saccharomyces (J. N. Stern et al., eds.), pp. 97-142. Cold Spring Harbor Laboratory, 1981.]
13.2.3. Регуляция размеров клетки зависит от факторов контроля клеточного цикла, действующих в точке старта
Скорость роста простых свободноживущих организмов, таких как дрожжи, зависит главным образом от поступления питательных веществ. В условиях нехватки пищи дочерние клетки при быстрых циклах клеточного деления становились бы чрезвычайно мелкими; поэтому клеткам необходим механизм, регулирующий скорость прохождения клеточного цикла и, в частности, хромосомного цикла в соответствии со скоростью роста клетки (рис. 13-20). Как осуществляется такая регуляция?
И синтез ДНК, и митоз - это сложные динамические процессы, которые трудно замедлить или прервать в условиях нехватки питательных веществ. У дрожжей, как и у большинства других эукариотических организмов, продолжительность этих фаз цикла остается более или менее постоянной, несмотря на большую изменчивость внешних условий. Вместо этого во время голодания обычно удлиняется фаза G1, хотя у делящихся дрожжей имеется также важный регулирующий механизм, называемый митотическим контролем, который действует в фазе G2.
Если продолжительность фазы G1 может меняться под влиянием внешних факторов, а S-фаза не изменяется, то в G1 должна существовать критическая точка, где начинается последовательность событий S-фазы, и внешние факторы уже не действуют на дальнейший ход клеточного цикла. Такую критическую точку называют точкой старта (Start). Для большинства эукариотических клеток точка старта (или эквивалентная ей точка рестрикции у клеток млекопитающих) отмечает момент перехода к безостановочному завершению цикла клеточного деления.
13.2.4. Клетки проходят через точку старта только после достижения критических размеров [14]
Для клетки почкующихся дрожжей в среде, бедной питательными веществами, фаза G1 является периодом медленного роста, когда хромосомный цикл, видимо, приостановлен; выход из G1 т. е. прохождение через точку старта, происходит лишь после того, как клетка достигнет некоторых стандартных размеров (рис. 13-20, Б). В более богатой среде G1 короче, но величина клетки при прохождении точки старта практически такая же; и если подобрать условия роста, при которых дочерние клетки после деления будут ненормально крупными или ненормально мелкими, то они соответственно сократят или продлят время своего пребывания в фазе G1 так, чтобы пройти через точку старта, имея стандартные размеры.
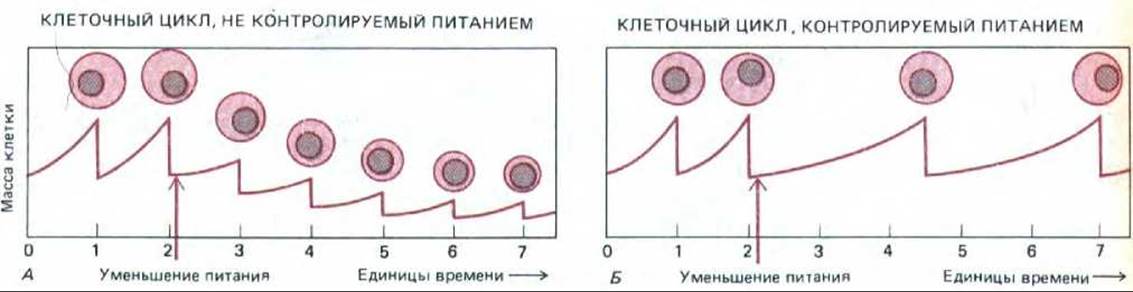
Рис. 13-20. Зависимость между скоростью роста, размерами клетки и циклом деления у свободноживущего организма, такого как дрожжи. А. Если при недостатке пищи клетки продолжают делиться с прежней скоростью, то дочерние клетки после каждого деления будут становиться все мельче до тех пор, пока масса каждой из дочерних клеток не сравняется с тем малым количеством вещества, которое синтезируется за время одного цикла. Б. Обычно при нехватке питательных веществ дрожжевые клетки замедляют темп деления: поскольку клетка не может пройти определенную точку цикла, не достигнув некоторых «стандартных» размеров, деление замедляется и величина клеток остается более или менее постоянной. (За единицу времени выбрана наблюдаемая длительность цикла при избытке питательных веществ.)
О том, как клетки «чувствуют» свою величину, мало что известно, хотя многие данные указывают на то, что какой-то механизм для этого существует. Например, если у растущей гигантской амебы Amoeba proteus многократно отрезать часть цитоплазмы, не позволяя таким клеткам достичь нормальных размеров, то она не будет делиться даже на протяжении нескольких недель, несмотря на энергичный рост, тогда как контрольная клетка делится примерно раз в сутки. Возможный намек на то, как клетка «ощущает» свои размеры, содержится в том факте, что величина эукариотической клетки обычно пропорциональна ее плоидности: диплоидная клетка в два раза больше гаплоидной, а тетраплоидная в два раза больше диплоидной (см. рис. 13-40 и 13-41). Можно предположить, что решающую роль играет отношение клеточного объема к числу копий какого-то гена (или набора генов) или к общему количеству ДНК (а не отношение, скажем, объема клетки к ее поверхности). Например, некая растворимая молекула М (допустим, какая-то РНК) могла бы непрерывно синтезироваться ДНК-зависимым способом; если М нестабильна с постоянным периодом полужизни, то общее количество М в каждой клетке будет постоянным и будет находиться в определенном соотношении с количеством ДНК. По мере увеличения объема клетки концентрация М будет снижаться; падение концентрации ниже некоторого критического уровня могло бы быть сигналом к прохождению точки старта.
Каков бы ни был этот механизм, прохождение через точку старта должно соответствовать скачку в состоянии какого-то молекулярного переключателя. Четыре гена cdc у почкующихся дрожжей и два гена у делящихся дрожжей действуют в точке старта или около нее, и они, возможно, кодируют компоненты такого регуляторного механизма. Клетки с термочувствительными мутациями этих генов не смогут вступить в хромосомный цикл и вырастут ненормально крупными, если повысить температуру до рестриктивного уровня, пока они еще не достигли критических размеров, чтобы пройти точку старта. Ниже мы расскажем подробнее об одном из старт-контролирующих генов (cdc28) у почкующихся дрожжей и соответствующем ему гене (cdc2) у делящихся дрожжей. Эти два гена примечательны одной своей дополнительной функцией, особенно выраженной у делящихся дрожжей: их продукты необходимы не только для прохождения точки старта, но и для второй контрольной точки цикла-начала митоза.
Далее мы увидим, что у клеток высших эукариот в фазе G1 имеется контрольная точка, аналогичная точке старта, хотя правила прохождения через эту точку сложнее, чем у дрожжей. При нарушении этих правил возникают раковые опухоли. По одной только этой причине гены, участвующие в механизме точки старта, представляют особый интерес.
13.2.5. Прохождение через точку старта зависит от протеинкиназы, родственной М-стимулирующему фактору (MPF) [15]
Дрожжи с их быстрым размножением и простой одноклеточной организацией - привлекательный объект для генной инженерии, и их легко использовать для включения ДНК, добавляемой в питательную среду. В принципе это позволяет клонировать нормальную («дикого типа») форму любого из генов cdc. Как показано на рис. 13-21, интересующий нас клон может быть без труда выделен благодаря его способности избавлять соответствующего мутанта cdc от его аномалий.
Такой подход был использован для клонирования гена cdc28 почкующихся дрожжей и гена cdc2 делящихся дрожжей; при этом было обнаружено несколько удивительных соответствий между этими двумя типами дрожжей и между дрожжами и позвоночными в отношении контроля точки старта и митоза. Гены cdc2 и cdc28 у дрожжей гомологичны и по последовательности нуклеотидов, и по функции: у делящихся дрожжей с мутацией гена cdc2 функциональный дефект можно устранить путем введения гена cdc28 от почкующихся дрожжей. Более того, у тех же мутантов по cdc2 сходного результата можно достичь, вводя им клонированный фрагмент ДНК человека, в которой, оказывается, тоже имеется последовательность, гомологичная cdc2/28. По-видимому, этот компонент механизма, управляющего клеточным циклом, является общим для дрожжей, млекопитающих и, вероятно, всех других эукариот.
Имея клонированный ген cdc2/28, сравнительно легко выделить белок, кодируемый этим геном. Он представляет собой протеинкиназу и, судя по всему спектру структурных и функциональных критериев, видимо, является дрожжевым гомологом киназной субъединицы MPF позвоночных. Кроме того, клонирование гена показало, что другой ген cdc делящихся дрожжей-cdc13, продукт которого взаимодействует с продуктом гена cdc2, в высокой степени гомологичен гену циклина (разд. 13.1.11). Эти данные показывают, что MPF и циклин, вероятно, имеют универсальное значение в клеточном цикле эукариот; а двойственная роль cdc2/28 у дрожжей - в начале М-фазы и в точке старта - позволяет предположить, что у позвоночных соответствующая субъединица MPF или близкая к ней молекула тоже может каким-то образом участвовать в контроле инициации цикла деления во время фазы G1.

Рис. 13-21. Метод выделения генов cdc из библиотеки ДНК. Клон редкой ДНК, содержащей нормальную копию дикого типа определенного гена cdc, легко отобрать, так как содержащая его плазмида делает соответствующую мутантную клетку неспособной расти при высокой температуре. И ген cdc2 делящихся дрожжей, и ген cdc28 почкующихся дрожжей первоначально были выделены таким способом. Повторяя эту процедуру с клонами кДНК из библиотеки ДНК человека в подходящей плазмиде, удалось выделить гены человека, способные заменять некоторые гены cdc дрожжей.
В самом деле, исследования на делящихся дрожжах показывают, что изменения в состоянии фосфорилирования этой регуляторной молекулы могут быть тем механизмом, с помощью которого клетки координируют свою готовность к началу цикла деления с условиями среды.
Заключение
Дрожжи - одноклеточные эукариотические организмы, очень подходящие для генетического анализа. И у почкующихся, и у делящихся дрожжей было идентифицировано множество мутаций, затрагивающих цикл клеточного деления (cdc), и клонированы соответствующие гены дикого типа. У дрожжевых и многих других эукариотических клеток, несмотря на изменчивые условия питания, поддерживаются стандартные размеры клетки с помощью механизма, который препятствует прохождению клетками критической точки (называемой точкой старта) и запускает цикл деления, когда они достигают пороговых размеров. У дрожжей некоторые из ключевых генов cdc, участвующих в этом контроле, были идентифицированы и их нуклеотидные последовательности определены. Один из них (обозначаемый cdc2 у делящихся дрожжей и cdc28-y почкующихся дрожжей) кодирует протеинкиназу, гомологичную MPF; другой ген (cdc!3) кодирует дрожжевой гомолог циклина.