Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Транспорт веществ в клетке
Вакуолярный транспорт
Гликопротеины, переносимые из шероховатого эндоплазматического ретикулума в другие места, "упаковываются" в мелкие транспортные пузырьки (везикулы), которые отщепляются от промежуточной части эндоплазматического ретикулума.
Стенка этих пузырьков состоит из липидов и белков мембраны эндоплазматического ретикулума, а сами пузырьки содержат растворимые гликопротеины, захваченные из полости эндоплазматического ретикулума.
Большинство из этих пузырьков сливается, образуя плоские цистерны на ближней к ядру стороне аппарата Гольджи, с так называемой формирующейся, или цис-стороной (формирующий полюс).
В цистернах аппарата Гольджи происходит дальнейшее "созревание" синтезированных в шероховатом эндоплазматическом ретикулуме и перешедших в аппарат Гольджи белков.
"Созревание" белков включает различные ковалентные модификации, в результате которых белки приобретают свою функциональноактивную структуру. Кроме того, очень сильно модифицируются связанные с остатком аспарагина олигосахариды, присоединившиеся ранее к белкам в эндоплазматическом ретикулуме - некоторые остатки сахаров избирательно отщепляются, а некоторые новые добавляются.
Процессинг олигосахаридов осуществляется с помощью нескольких различных чрезвычайно сложных и точно "запрограммированных" ферментных механизмов. Выбор "программы" модификации определяется каким-то (пока неизвестным) свойством каждой индивидуальной полипептидной цепи.
Ферменты, осуществляющие модификацию белков, как и сами модифицирующиеся белки, тоже, вероятно, попадают в аппарат Гольджи из эндоплазматического ретикулума. В настоящее время считается, что после процессинга присоединенный к белку олигосахарид становится "адресом", по которому должен быть доставлен данный белок.
"Созревшие" белки снова "упаковываются" в транспортные пузырьки, но уже на зрелой, или транс-стороне (декретирующий полюс).
Обобщенная схема везикулярного транспорта представлена на рисунке 34. Белки, синтезированные в шероховатом эндоплазматическом ретикулуме (рисунок 34(1)) упаковываются в транспортные везикулы (рисунок 34(2)), которые отсоединяются от шероховатого эндоплазматического ретикулума и сливаются вместе, образуя новую цистерну цис-Гольджи сети (cis-Golgi network, CGN). Ферменты и белки эндоплазматического ретикулума, которые попали в такой пузырек, а также мембранные белки, которые обеспечивают формирование везикул, возвращаются в шероховатый ЭР для "повторного" использования (рисунок 34(3)) в пузырьках, которые отъединяются от цис-стороны аппарата Гольджи и сливаются с мембраной шероховатого эндоплазматического ретикулума.
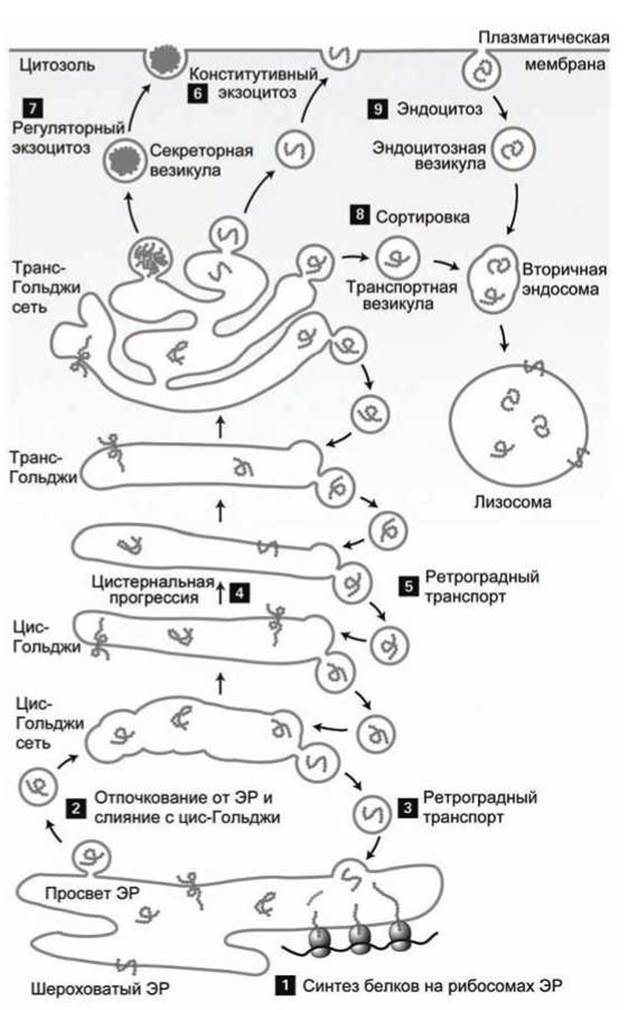
Рисунок 34 - Обобщенная схема везикулярного транспорта
Каждая цистерна Гольджи из цис-области физически перемещается через промежуточную область в транс-область аппарата Гольджи посредством невезикулярного процесса, который называется цистернальная прогрессия (cisternalprogression) (рисунок 34(4)).
"Возврат" специфических для каждой области аппарата Гольджи белков осуществляется везикулярным механизмом (рисунок 34(5)). Во всех клетках определенные белки постоянно перемещаются к поверхности в везикулах, отъединившихся от транс-стороны аппарата Гольджи, обеспечивая постоянную секрецию (рисунок 34(6)) белков за пределы клетки (конститутивный экзоцитоз). В некоторых типах клеток определенные белки "запасаются" в секреторных везикулах (рисунок 34(7)) и секретируются только после получения клеткой внешнего нейро- или гормонального сигнала (регуляторный экзоцитоз)
Белки и липиды, которые предназначены для лизосом (рисунок 34(8)), отсоединяются от транс-стороны аппарата Г ольджи и присоединяются к вторичным эндосомам, которые затем сливаются с лизосомой. Белки из внеклеточного пространства и мембранные белки плазматической мембраны клетки, которые в процессе эндоцитоза (рисунок 34(9)) формируют эндоцитозные везикулы (отъединяющиеся от плазматической мембраны внутрь клетки), также перемещаются внутрь лизосом через эндосомы.
Окаймлённые пузырьки. Большинство транспортных пузырьков- везикул представляют особый класс внутриклеточных пузырьков, которые на микрофотографиях выглядят окруженными со стороны цитоплазмы щетинообразной каймой, и потому называются окаймленными пузырьками. Диаметр окаймленных пузырьков варьирует от 50 до 250 нм.
Окаймление образуют три типа белков:
1) СОРП-белок покрывает везикулы, транспортирующие белки из шероховатого эндоплазматического ретикулума в аппарат Гольджи,
2) COPI-белок покрывает везикулы, осуществляющие "возвратный" транспорт белков между цистернами Гольджи и в шероховатый эндоплазматический ретикулум,
3) клатрин окаймляет везикулы, которые транспортируют белки от плазматической мембраны и от транс-Гольджи сети к вторичным эндосомам (рисунок 35).
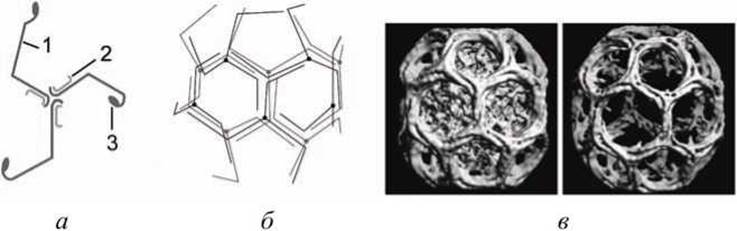
Рисунок 35 - Структура клатринового покрытия: а - трискелион; б - упаковка трискелионов в клатриновую оболочку; в - клатриновые корзинки, образованные спонтанной агрегацией трискелионов in vitro; 1 - клатрин; 2 - малый полипептид; 3 - узел связывания клатринов при агрегации
Клатрин (180 кДа) вместе с меньшим полипептидом (35 кДа) образует характерный многогранный чехол на поверхности окаймленных пузырьков.
Основным структурным элементом чехла служит белковый комплекс трискелион, состоящий из трех полипептидных цепей клатрина и трех малых (35 кДа) полипептидов.
Трискелионы образуют на поверхности окаймленных пузырьков корзиноподобные сетки из шестиугольников и пятиугольников. Обособленные трискелионы при соответствующих условиях способны к спонтанной агрегации. При этом даже в отсутствие пузырьков формируются типичные многогранные корзинки (рисунок 35(b)).
В мембраны везикул интегрированы множество вспомогательных белков. Эти белки, во-первых, в процессе формирования пузырька "вылавливают" из цистерн аппарата Гольджи белки с одинаковыми олиго-сахаридными "адресами", и, во-вторых, узнают компоненты мембраны, с которой должен слиться данный пузырек (рисунок 36).
Известно, что содержимое каждого транспортного пузырька доставляется точно по правильному "адресу" к определенной внутриклеточной мембране, поэтому должны существовать различные субпопуляции транспортных пузырьков, имеющих на своей поверхности уникальные вспомогательные белки, так называемые маркеры стыковки (v-SNARE белки) (рисунки 36 и 37), которые опознаются комплементарными рецепторами (t-SNARE белки) на мембранах мишеней и формируют SNARE-комплекс.

Рисунок 36 - Схема везикулярных процессов: а - формирование окаймленного пузырька из донорной мембраны, б - слияние везикулы с мебраной-целью. 1 - донорная мембрана; 2 - водорастворимый транспортируемый белок; 3 - ГТФ-связывающий белок; 4 - v-SNARE-белок; 5 - интегрированный в мембрану транспортируемый белок; 6 - мембранный рецептор водорастворимых транспортируемых белков; 7 - окаймляющие белки; 8 - мембрана-цель; 9 - t-SNARE-белки; 10 - t-SNARE-комплекс
Окаймление играет ключевую роль в формировании везикулы, но вскоре после отделения везикулы окаймляющие белки диссоциируют с её поверхности (потеря каймы), и отсутствие окаймления позволяет везикуле слиться с мембраной в конце транспортировки. Схема везикулярного транспорта между ШЭР и цис-Гольджи представлена на рисунке 37.
Прямой (антероградный) транспорт обеспечивается СОРП-окаймленными везикулами (рисунок 37(1,2,3)) - которые образуются полимеризацией СОРП-белков на поверхности мембраны эндоплазматического ретикулума.

Рисунок 37 - Везикулярный транспорт белков между эндоплазматическим ретикулумом и цис-Гольджи-областью
Диссоциация СОРП-белков с поверхности везикулы освобождает v-SNARE-белки. Спаривание v-SNARE с t-SNARE-белками на поверхности цис-Гольджи сети образует SNARE-комплексы, в результате чего становится возможным слияние мембран.
Обратный (энтероградный) транспорт (рисунок 37(4,5,6)), осуществляемый с помощью окаймления везикул СОРІ-белками, обеспечивает возврат (рециклирование) мембранных липидов, специфических белков (например, v-SNARE) и ошибочно захваченных белков эндоплазматического ретикулума из цис-Гольджи сети обратно в ШЭР.
Транспортные пузырьки, сливающиеся с плазматической мембраной и высвобождающие свое содержимое в межклеточное пространство, называются секреторными, или экзоцитозными пузырьками. Наряду с окаймленными пузырьками в клетке образуются и окаймленные ямки, т.е. эндоцитозные пузырьки, отпочковывающиеся внутрь клетки от окаймленных участков плазматической мембраны.
Когда клетке нужно "выловить" из межклеточной жидкости какое- либо вещество, она, как правило, использует механизм опосредуемого рецепторами эндоцытоза, или адсорбционный эндоцитоз, при котором клетка производит белки-рецепторы для этого вещества, которые встраиваются в состав окаймленных участков плазматической мембраны. Все молекулы, связавшиеся с рецепторами, быстро проникают внутрь клетки, поскольку окаймленные ямки постоянно отщепляются внутрь, образуя окаймленные пузырьки.
Эти пузырьки быстро теряют свою кайму и сливаются с другими пузырьками, образуя более крупные пузырьки, называемые эндосомами. Содержимое эндосом может использоваться клеткой. Хорошо изученный пример использования адсорбционного эндоцитоза - это поглощение животными клетками холестерола из внеклеточной среды.
Некоторые транспортные пузырьки, которые отпочковываются от самой крайней цистерны транс-стороны аппарата Гольджи, становятся первичными лизосомами. Гликопротеины, которыми загружены эти пузырьки, имеют олигосахарид с фосфорилированным остатком маннозы (рисунок 38). Этот маннозо-6-фосфатный маркер и определяет судьбу таких пузырьков. После отщепления маннозо-6-фосфатного маркера гликопротеины первичных лизосом становятся активными гидролитическими ферментами (т. е. расщепляющими ферментами, осуществляющими перенос функциональных групп на молекулу воды).
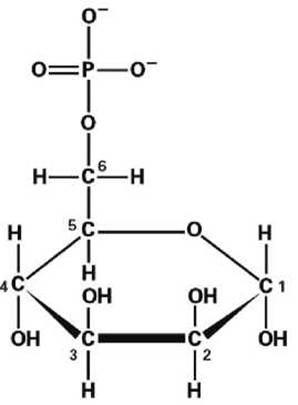
Рисунок 38 - Манноза-6-фосфат
Лизосомы расщепляют до мономерных компонентов макромолекулы, захваченные клеткой в процессе эндоцитоза, или отдельные участки цитоплазмы и органоиды, утратившие свое значение в результате старения или использующиеся как материал для поддержания жизнедеятельности клетки в неблагоприятных экстремальных условиях.
В первом случае первичная лизосома сливается с эндоцитозным пузырьком. Во втором случае подлежащий "перевариванию" участок цитоплазмы или органоид сначала окружается мембраной, а затем получившийся пузырек сливается с первичной лизосомой.
Первичная лизосома, слившаяся с пузырьком, содержащим субстрат для расщепления, называется уже вторичной лизосомой. Мономерные продукты расщепления транспортируются из лизосомы в цитоплазму и могут использоваться клеткой.
Сейчас известно около 60 гидролитических ферментов, содержащихся в лизосомах, которые способны разрушать практически все природные полимерные органические соединения. Все они обладают наибольшей активностью при pH ≈ 5. Именно такое значение pH поддерживается внутри лизосомы встроенными в их мембрану протонными АТФазами (протонными помпами), использующими энергию АТФ для накачивания ионов Н+ в полость этих органелл. Хотя в нормальных условиях мембрана лизосомы непроницаема для гидролитических ферментов, необходимость кислой среды для их активной работы защищает цитоплазму клетки от разрушения при возможной "утечке" ферментов.