Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Фотосинтетические системы
Разделение зарядов в реакционном центре фотосистемы
Когда молекула хлорофилла а, расположенная в составе "специальной пары" - димера хлорофиллов - вблизи поверхности реакционного центра, обращенной к люмену тилакоида, поглощает свет в максимуме поглощения 680 нм, эта молекула хлорофилла а переходит из основного энергетического состояния в первое возбужденное состояние и её энергия увеличивается на 42 ккал/моль. Реакционный центр устроен таким образом, что возбужденный электрон за счет квантово-механического туннелирования уходит с молекулы хлорофилла ещё до того момента времени, когда он смог бы релаксировать обратно в основное состояние.
Процесс туннелирования внутри реакционного центра по цепи промежуточных акцепторов электрона заканчивается на поверхности мембраны тилакоида, обращенной к строме на первичном акцепторе электрона - на молекуле пластохинон Q (рисунок 180(6)), которая подобна молекуле убихинона (рисунок 180(a)).
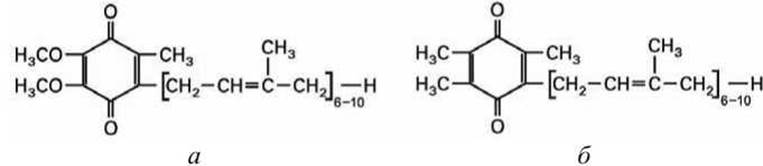
Рисунок 180 - Липофильные переносчики электрона: а - убихинон в дыхательной цепи митохондрии; б - пластохинон в фотосинтетической электротранспортной цепи хлоропласта
Такой перенос электронов по цепи промежуточных акцепторов, стимулированный поглощением фотона, называется фотоэлектронным транспортом.
Особенности фотоэлектронного транспорта зависят от особенностей белкового окружения, как хлорофилла, так и акцепторов электрона в реакционном центре. В результате фотоэлектронного транспорта вблизи люминальной поверхности реакционного центра остается положительно заряженная молекула хлорофилла а+, а вблизи поверхности, обращенной к строме, образуется отрицательно заряженный акцептор (Q-) (рисунок 181). Тем самым, в реакционном центре реализуется фотосинтетическое разделение зарядов (charge separation).
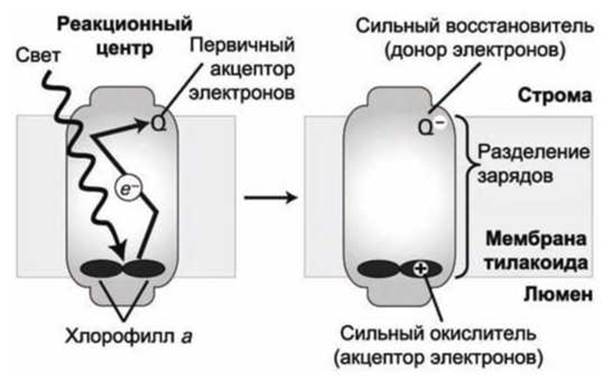
Рисунок 181 - Фотоэлектронный транспорт в реакционном центре
Пластохинон Q является сильным восстановителем, который эффективно передает избыточный электрон другим молекулам, прежде всего, молекуле НАДФ.
Положительно заряженный хлорофилл а+ является сильным окислителем, захватывающим электрон на люминальной поверхности с целью регенерации исходного состояния хлорофилла а.
В растениях такой захват электронов четырьмя молекулами хлорофилл а+ в итоге приводит к удалению четырех электронов из двух молекул воды, связанных с люминальной поверхностью реакционного центра с образованием молекулярного кислорода:
![]()
Эти оба сильных биологических восстановителя и окислителя являются источником энергии для всех последующих реакций фотосинтеза — для переноса электронов, синтеза АТФ и фиксации СО2.
Хлорофилл а поглощает фотоны и с длинами волн меньше, чем 680 нм (более высокоэнергетичные фотоны) (рисунок 38). В результате такого поглощения молекула возбуждается в одно из более высоких возбужденных состояний. Однако эти возбужденные состояния всё равно релаксируют в нижайшее (первое) возбужденное состояние за очень короткое время порядка одной пикосекунды (10-12 с), а избыточная энергия при этом выделяется в виде тепла.
Поскольку процессы фотоэлектронного транспорта, приводящего к разделению зарядов, начинаются всегда только с первого возбужденного состояния реакционного центра хлорофилла а, то квантовый выход - число актов фотосинтеза приходящееся на один поглощенный фотон - не зависит от длины волны света (с длинами волн короче (а, соответственно, с энергиями выше) чем 680 нм).