Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Молекулярные механизмы фотосинтеза
Фотосистема пурпурных бактерий
Молекулярный кислород в результате фотосинтеза образуется в цианобактериях, водорослях и растениях, но не образуется в зеленых и пурпурных бактериях. В последних существуют только фотосистемы типа І, ФС1 (photosystem I, PSI), в которых происходит восстановление НАДФ+ до НАДФН, а в цианобактериях, водорослях и растениях дополнительно есть ещё фотосистемы типа II, ФС2 (photosystem II, PSU), в которых образуется О2 из Н2О.
Механизм разделения зарядов в фотосистеме пурпурных бактерий идентичен описанному выше механизму разделения зарядов в фотосистемах растений. Свет поглощает так называемая "специальная пара" - пигментный димер молекул бактериохлорофиллов а, очень быстро (за время менее 4 пикосекунд) возбужденный электрон переносится на молекулу феофитина (pheophytin, Ph), а хлорофилловый димер остается положительно заряженным.
Затем за 200 пс электрон переносится на молекулу менахинона QA, и, наконец, за 200 мкс (самый медленный этап) происходит перенос электрона на молекулу убихинона QB (рисунок 183) и образование полухинона QB- (см. рисунок 166).
Поглощение второго фотона этим же реакционным центром добавляет второй электрон на этот полухинон, в результате чего образуется полностью восстановленный хинон (дигидрохинон, QH2, (рисунок 166)), который высвобождается из реакционного центра и диффундирует по плазматической мембране бактерии к Q0-центру на экзоплазматической стороне комплекса цитохром bс1.
Связывание хинона QH2 с Q0-центром приводит к высвобождению двух протонов в периплазматическое пространство (пространство между плазматической мембраной и клеточной стенкой бактерии). Таким образом, поглощение света приводит к выкачиванию протонов из цитозоля и генерации протондвижущей силы на плазматической мембране бактерии.
Одновременно, хинон QH2 отдает два электрона, которые переносятся внутри комплекса цитохром bс1, совершенно аналогично тому, как это происходит в митохондриальном комплексе CoQH2-цитoxpoм с редуктаза (рисунок 167). Q-цикл в комплексе цитохром bс1 бактерии, так же как и Q-цикл в митохондрии, выкачивает ещё один дополнительный протон из цитозоля в межмембранное (периплазматическое) пространство, тем самым ещё более увеличивая протондвижущую силу.
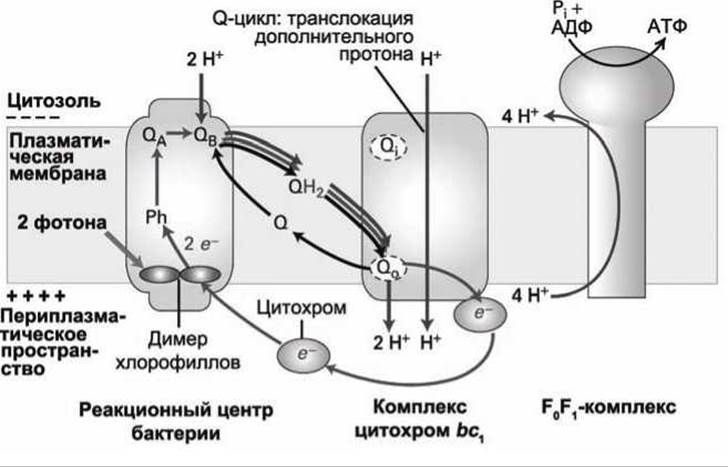
Рисунок 183 - Цепь переноса электронов в фотосистеме пурпурных бактерий
Акцептором электронов, прошедших через комплекс цитохром bс1, является малый водорастворимый железосодержащий белок-переносчик цитохром, который находится в периплазматическом пространстве бактерии. Получив электрон, ион железа в цитохроме изменяет заряд (восстанавливается) с Fe3+ на Fe2+.
Восстановленный цитохром (аналогично цитохрому с в митохондриях (рисунок 167)) диссоциирует с комплекса цитохром bc1 и диффундирует к реакционному центру, где и замыкается Q-цикл — электрон переходит на положительно заряженный хлорофилл а+, возвращая в исходное состояние и хлорофилл, и ион железа цитохрома.
В результате такой циклической (периодической) циркуляции электрона по замкнутой цепи не образуется кислород и не происходит восстановления коферментов.
Возможно также альтернативное использование электронной цепи фотосистем бактерий, при котором электроны движутся по этой цепи однонаправлено (линейно), а не циклически. В этом случае электроны, возбужденные в хлорофиллах реакционных центров, переносятся на кофермент НАД⊕ (а не на НАДФ+, как в растениях), восстанавливая его до НАДН. Окисленный хлорофилл а+ восстанавливается электроном, который приносит цитохром с. А вот для восстановления цитохрома с бактерии используют электроны либо из сероводорода H2S (образуя атомарную серу S), либо просто из молекулярного водорода Н2.
Ни в том, ни в другом случае не используется вода Н2O в качестве донора электронов, поэтому и в этом, альтернативном, случае "линейного" использования цепи переноса электронов фотосистемой бактерии молекулярный кислород O2 не образуется.
Протондвижущая сила создается в обоих случаях - и в случае циклического переноса, и в случае линейного переноса электронов в бактериальных фотосистемах. Так же, как и в других организмах эта протондвижущая сила используется расположенными на плазматической мембране белками: АТФ-синтазами для синтеза АТФ, и белками- переносчиками для транспорта молекул против градиента концентрации.