Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Аллели и мутации
Сегрегация мутаций при селекционировании
Доминантность или рецессивность аллелей определяют с помощью жизненного цикла организмов.
Различают два типа деления клеток - митоз и мейоз.
Большинство клеток (соматические клетки) организма делится митотически.
Мейоз характерен для зародышевых клеток, из которых в результате мейоза образуются гаметы (у животных их называют сперматозоидами и яйцеклетками). Подобно соматическим клеткам, премейотические зародышевые (эмбриональные) клетки являются диплоидными, содержащими два гомолога каждого морфологического типа хромосом.
Два гомолога, образующих каждую пару гомологических хромосом унаследованы от разных родителей и, поэтому, их гены могут существовать в различных аллельных формах.
Основные этапы митоза и мейоза представлены на рисунке 74.
В обоих случаях, и для соматической клетки, и для премейотической клетки эмбриона, исходная клетка имеет две копии каждой хромосомы (2n), одну материнскую и одну отцовскую.
При митозе реплицированные хромосомы, каждая состоящая из двух сестринских хроматид, выстраиваются в центре клетки так, чтобы обе дочерние клетки получили материнский и отцовский гомологи каждой хромосомы.
При мейозе в ходе первого деления, каждая реплицированная хромосома спаривается (выстраивается парами) с её гомологичным партнером в центре клетки - такое разделение на пары называется синопсисом. Затем обе реплицированные хромосомы (из пары) при делении №1 попадают в разные дочерние клетки. А эти дочерние клетки, в свою очередь,
второй раз делятся (деление №2), но без предварительной репликации ДНК. При этом каждая клетка получает по одной хроматиде-"сестре". Таким образом, каждая диплоидная клетка в результате мейоза даёт четыре гаплоидные (ln) клетки (гаметы).
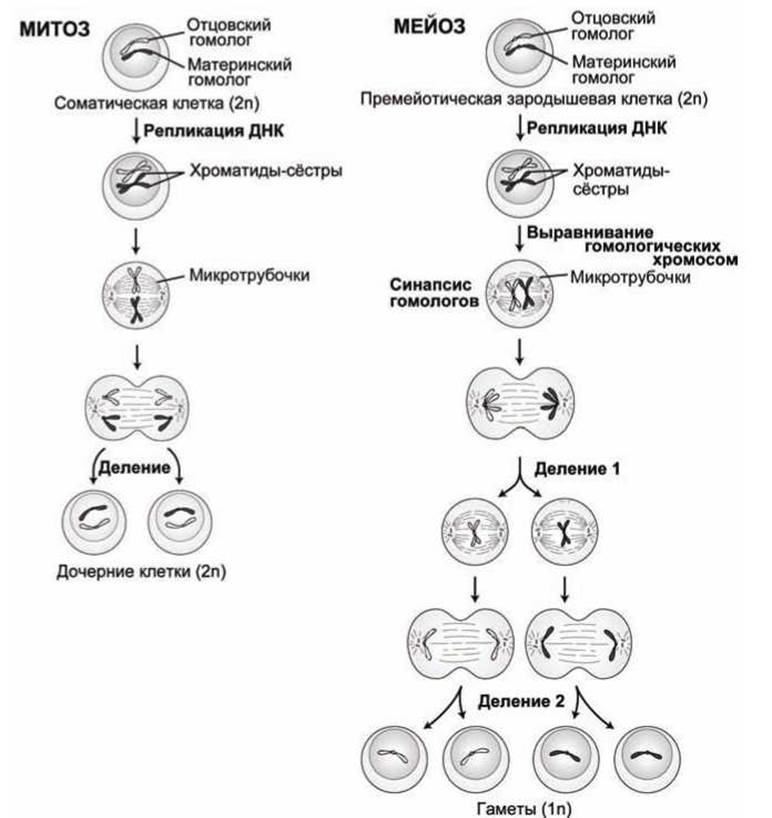
Рисунок 74 - Этапы митоза и мейоза
При митозе за репликацией ДНК всегда следует деление клетки и образуется две диплоидные дочерние клетки.
При мейозе после одной репликации ДНК следует два различных деления клетки, в результате которых и образуются четыре гаплоидных (ln) клетки, каждая из которых содержит только одну хромосому из каждой гомологичной пары.
Такое пропорциональное разделение (или сегрегация) реплицированных хромосом по дочерним клеткам в ходе первого мейотического деления является произвольным, что приводит к образованию дочерних клеток со случайным сочетанием отцовских и материнских хромосом.
Чтобы не усложнять эксперимент, обычно ограничиваются селекционированием гомозиготных по исследуемому гену штаммов. В таких "истинных селекционируемых" штаммах (true-breeding strains) каждая особь будет получать одинаковую аллель от каждого из предков и, поэтому, набор аллелей не будет изменяться от одного поколения к следующему.
Когда "истинный" (гомозиготный) мутантный штамм спаривается с "истинным" "диким" (wild) штаммом первое дочернее потомство (F1) будет гетерозиготным (рисунок 75).
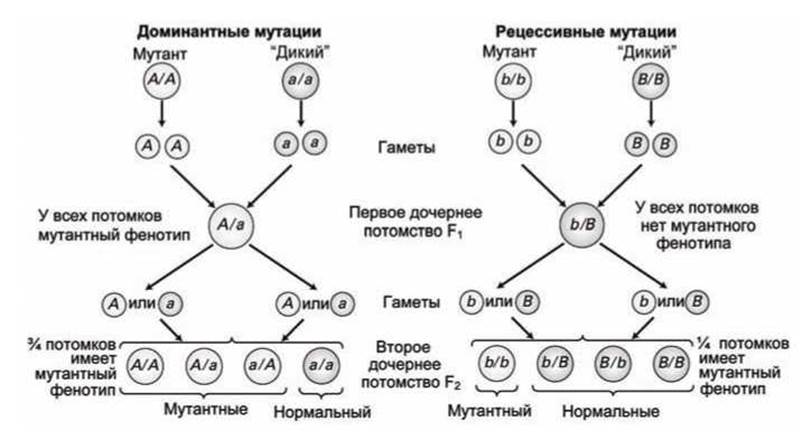
Рисунок 75 - Сегрегация доминантных и рецессивных мутаций
Если F1 потомство демонстрирует мутантный фенотип, то это означает, что исходный мутант доминантный, если же F1 потомство демонстрирует "дикий" фенотип, то, значит, исходный мутант рецессивный.
Когда скрещиваются гетерозиготные F1 особи с доминантной мутантной аллелью, то три четверти их F2 потомства будет проявлять мутантные особенности, в то время как при скрещивании гетерозиготных F1 особей с рецессивной мутантной аллелью, только одна четверть их F2 потомства будет проявлять мутантные особенности.
Дрожжи Saccharomyces существуют как в гаплоидной, так и в диплоидной формах.
В этих одноклеточных эукариотах скрещивание между гаплоидными клетками позволяет определить доминантность или рецессивность мутантных аллелей. Гаплоидные клетки дрожжей, содержащие однократный набор хромосом, могут быть в одной из двух форм, обозначаемых a и а (рисунок 76).
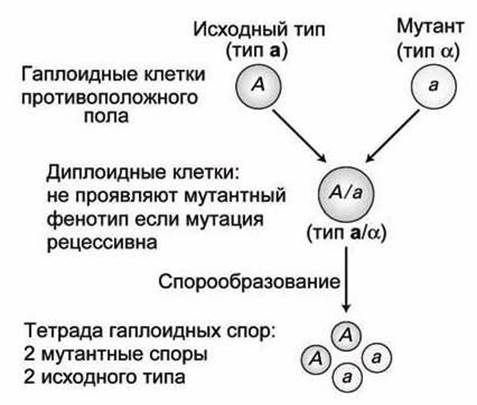
Рисунок 76 - Сегрегация аллелей в дрожжах
Гаплоидные клетки этих двух противоположных типов могут быть скрещены для получения а/а диплоидов, которые являются гетерозиготными по мутантной аллели.
Если эти диплоиды проявляют мутантные свойства, значит мутантная аллель является доминантной, но если диплоиды проявляются как исходный дикий тип, значит мутантная аллель рецессивна.
Когда а/а диплоид испытывает недостаток пищи (голодание), клетка претерпевает мейоз, в результате чего образуется тетрада из четырёх гаплоидных спор, две типа а, и две типа а. Споруляция (спорообразование) гетерозиготной диплоидной клетки порождает две споры, содержащие мутантную аллель и две, несущие исходную аллель дикого типа (рисунок 76).
При благоприятных условиях споры дрожжей оживают, порождая вегетативные гаплоидные штаммы обоих типов.