РОСТ И КУЛЬТИВИРОВАНИЕ БИООБЪЕКТОВ - В. М. Самыгин - 2016
ГЛАВА 5. РОСТ И РАЗМНОЖЕНИЕ МИКРООРГАНИЗМОВ
Постоянно осуществляемый микроорганизмами синтез клеточных компонентов стимулирует рост клеток и последующее их размножение. Рост клеток - это процесс увеличения размеров, объема, массы каждой отдельной особи в ходе индивидуального развития, обусловленный синтезом клеточного материала. Рост начинается после деления клетки, которая быстро достигает стадии зрелости, а затем приступает к размножению или переходит в стадию покоя. Рост характерен для всех групп микроорганизмов, кроме вирусов. Его изучают на изолированных клетках.
Размножение микроорганизмов - процесс самовоспроизведения, обеспечивающий сохранение вида. Способы и скорость размножения обусловлены геномом микроба и соответствием условий обитания его генетическим потребностям. Способы размножения у микроорганизмов разнообразны: бинарное деление - у большинства бактерий, почкование, спорообразование, половой процесс и др. - у грибов и дрожжей, особые способы размножения - у простейших (множественное деление, половое и др.).
При бинарном делении, инициатором которого является кольцевая ДНК бактерии, прикрепленная к определенному участку ЦПМ, из одной бактериальной клетки без предварительного обмена генетической информацией образуются две равноценные дочерние особи. Репликация ДНК наступает в определенной фазе роста под контролем гена-регулятора и гена- репликатора. Вновь образованная ДНК оказывается прикрепленной к соседнему участку ЦПМ. Между участками прикрепления исходной и новообразованной ДНК строго по экватору формируется перетяжка (у грамотрицательных бактерий) или перегородка (у грамположительных бактерий). После разделения дочерние клетки расходятся или формируют колониальные организмы в виде цепочек, гроздьев, пакетов и др.
От роста клеток следует отличать рост популяции - необратимое увеличение количества живого вещества (биомассы), обусловленное увеличением клеточной массы и количества клеток в единице объема питательной среды. Результаты количественного изучения роста могут быть представлены более информативно и точно, если анализируются различные параметры роста: удельная скорость роста, лаг-период, экономический коэффициент, метаболический коэффициент, урожай биомассы и др.
5.1. Кривая роста микроорганизмов в периодических условиях
Для роста биомассы в культуре необходимы следующие условия:
а) жизнеспособность засева;
б) наличие источника энергии;
в) внесение пищевых добавок, содержащих все компоненты, необходимые для синтеза биомассы;
г) отсутствие в среде ингибиторов, подавляющих рост клеток;
д) поддержание в среде подходящих физико-химических условий.
При внесении бактерий в питательную среду они обычно растут до
тех пор, пока содержание какого-нибудь из необходимых им компонентов среды не достигнет минимума, после чего рост прекращается. Если на протяжении этого времени не добавлять питательных веществ и не удалять конечных продуктов обмена, то получим так называемую периодическую культуру (популяцию клеток в ограниченном жизненном пространстве). Рост в такой «закрытой системе» подчиняется закономерностям, действительным как для одноклеточных, так и для многоклеточных организмов.
Кривая, описывающая зависимость логарифма числа живых клеток от времени, называется кривой роста. Типичная кривая роста имеет S-образную форму и позволяет различать несколько фаз роста, сменяющих друг друга в определенной последовательности. Она включает начальную (или лаг-) фазу; фазу ускорения роста; экспоненциальную (или логарифмическую), стационарную фазу; фазу замедления роста и фазу отмирания (рис. 5).
Рис. 5. Кривая роста микроорганизмов при периодическом культивировании: I - лаг-фаза; II - фаза ускорения роста; III - фаза экспоненциального роста; IV - фаза замедления роста; V -стационарная фаза; VI - фаза отмирания культуры
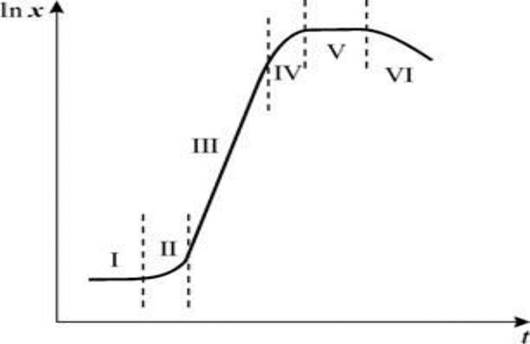
Начальная фаза. Эта фаза начинается с момента посева материала (внесения инокулята) до достижения культурой максимальной скорости роста. Продолжительность этой фазы зависит главным образом от объема и возраста инокулята, предшествовавших условий культивирования, а также от степени пригодности для роста выбранной среды. Внесение небольшого объема инокулята в большой объем свежей среды может привести к диффузии из клеток витаминов, кофакторов и ионов, которые необходимы для поддержания активности многих внутриклеточных ферментов. С увеличением возраста инокулята и при переносе клеток из бедной среды в богатую лаг-фаза удлиняется.
Если в качестве инокулята использована старая культура (в стационарной фазе роста), то клеткам придется сначала приспособиться к новым условиям путем синтеза РНК, образования рибосом и синтеза ферментов. Если в новой среде источники энергии и углерода отличаются от тех, что были в предшествующей культуре, то приспособление (адаптация) к новым условиям может быть связана с синтезом новых ферментов, которые ранее не были нужны и поэтому не синтезировались. Образование новых, адаптивных ферментов индуцируется новым субстратом. Причем время, необходимое для синтеза оптимального количества ферментов, может составлять от 10 мин до нескольких часов. У некоторых видов организмов индукция ферментов для использования новых источников углерода и энергии не происходит до тех пор, пока в среде есть хоть малые количества первоначального источника углерода и энергии.
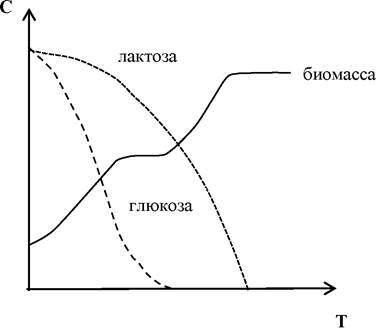
Наглядным примером влияния субстрата на синтез ферментов служит так называемая диауксия. Это явление двухфазного роста или двойного цикла роста наблюдается на средах, содержащих смесь питательных веществ. Например, из смеси глюкозы и лактозы Е. coli поглощает в первую очередь глюкозу. Глюкоза индуцирует в клетках сначала синтез ферментов, которые нужны для ее использования, и одновременно репрессирует (подавляет) синтез ферментов, необходимых для использования лактозы. Эти последние ферменты образуются лишь после того, как вся глюкоза будет израсходована. Такие регуляторные процессы объясняют наличие двух начальных фаз (рис. 6).
Рис. 6. Рост Е. coli в среде с различными сахарами (диауксия)
При оценке роста могут наблюдаться определенные отклонения от идеальной формы кривой роста. Если бактериальную массу измеряют по мутности, то часто наступлению стационарной фазы предшествует небольшое уменьшение оптической плотности. Это может быть артефактом, вызванным изменением отношения биомассы к оптической плотности. Другим отклонением является более быстрое увеличение числа организмов во время первого удвоения, что может быть обусловлено синхронным делением популяции одноклеточных организмов. Третий вариант - диауксия.
Количественное изменение состава бактериальной клетки во время начальной фазы роста сильнее всего затрагивает РНК: ее содержание повышается в 8-12 раз. Это указывает на участие РНК и рибосом (состоящих из РНК и белка) в синтезе ферментных белков.
Лаг-период может быть связан с инактивацией некоторых ингибиторов, присутствующих в среде. Такими ингибиторами иногда являются микроэлементы. Их токсическое действие связано с возможным образованием комплексов между входящими в состав питательной среды аминокислотами и ионами металлов. Токсическим свойством обладают органические кислоты, при этом с увеличением рН токсичность органических кислот уменьшается. Как ингибитор может вести себя субстрат, и тогда лаг-период будет зависеть от времени, необходимого для уменьшения концентрации субстрата до значений, при которых возможна максимальная скорость роста. Ингибирующее влияние на рост могут оказывать продукты метаболизма в инокуляте. Например, некоторые представители Lactobacillaceae при пересеве из анаэробной среды в аэробную могут накапливать перекись водорода, являющуюся сильным ингибитором роста. Перекись водорода индуцирует образование пероксидазы, которая удаляет перекись водорода. Лаг-период, обусловленный перекисью водорода, может увеличиваться с увеличением размера инокулята, поскольку образование перекиси водорода пропорционально концентрации клеток. Если инокулят состоит из споровых форм, то лаг-период, предшествующий вегетативному росту, соответствует времени прорастания спор.
Таким образом, существует множество причин, вызывающих задержку роста. Для того чтобы избежать или свести к минимуму лаг-период, инокулят следует брать из культуры, находящейся в конце логарифмической фазы, в условиях, как можно более близких к среде, используемой для пересева. Размер инокулята обычно следует брать по возможности больше.
Экспоненциальная (логарифмическая) фаза роста характеризуется постоянной максимальной скоростью деления клеток, которая зависит от вида бактерий и питательной среды. Энтеробактерии делятся через каждые 15-30 мин, у многих почвенных видов оно достигает 60-150 мин, а у Nitrobacter - даже 5-10 ч. Величина клеток и содержание в них белка у многих бактерий тоже остаются в экспоненциальной фазе постоянными. То есть в этом случае бактериальная культура состоит из «стандартных клеток». Если число клеток, содержание в них белка и их сухая биомасса увеличиваются с одинаковой скоростью, то за ростом культуры можно следить по одному из этих показателей.
Поскольку в экспоненциальной фазе скорость деления клеток относительно постоянна, эта фаза наиболее удобна для определения скорости деления клеток (скорости роста), изучения влияние факторов среды (рН, окислительно-восстановительного потенциала, температуры, аэрации и т. д.). Кроме того, в этой фазе определяют пригодность различных субстратов, а также следят за увеличением числа клеток или за мутностью (экстинкцией) клеточной суспензии.
Стационарная фаза наступает, когда число клеток перестает увеличиваться. Скорость роста зависит от концентрации субстрата - при уменьшении этой концентрации, еще до полного использования субстрата, она начинает снижаться. Поэтому переход от экспоненциальной фазы к стационарной происходит постепенно. Скорость роста может снижаться не только из-за нехватки субстрата, но также из-за большой плотности бактериальной популяции, низкого парциального давления О2 или накопления токсичных продуктов обмена; все эти факторы вызывают переход к стационарной фазе.
В ранней стационарной фазе размер бактериальной клетки достигает минимума. В поздней стационарной фазе (а также в фазе отмирания) часто наблюдаются искривленные или разбухшие клетки, называемые «инволюционными формами». Это связано либо с повреждениями клеток литическими ферментами клеточных стенок или плазматических мембран, либо слабой регуляцией синтеза клеточных компонентов. У грамположительных бактерий в стационарной фазе обычно наблюдается потеря способности клеток окрашиваться по Граму. Сопротивляемость бактерий физическим и химическим стрессам (влиянию гипотонической среды, внезапной смене температур) в стационарной фазе выше, чем в логарифмической. Для некоторых бактерий в стационарной фазе характерно образование спор.
В конце логарифмической фазы наблюдается синтез первичных метаболитов, то есть продуктов, необходимых для роста и выживания. В стационарной фазе синтезируются вторичные метаболиты - продукты, которые не требуются для роста и не существенны для выживания, однако они выполняют полезные функции и часто защищают от действия других конкурирующих микроорганизмов либо подавляют их рост (например, антибиотики). Количество биомассы, достигнутое в стационарной фазе, называют выходом или урожаем. Урожай зависит от природы и количества используемых питательных веществ, а также от условий культивирования.
Фаза отмирания. Гибель клеток во многом является результатом исчерпания запасов энергии в клетке. Клетки перестают делиться, гибнут и распадаются. Количество живых клеток резко уменьшается. Скорость отмирания бактерий варьирует в зависимости от условий и особенностей организма.
5.2. Параметры кривой роста
Основные параметры роста простой периодической культуры микроорганизмов (удельную скорость роста, время генерации, число клеточных делений) определяют в течение экспоненциальной фазы, тогда как максимальная концентрация жизнеспособных клеток в популяции характерна для стационарной фазы.
5.2.1. Удельная скорость роста
Если для роста биомассы в культуре соблюдены необходимые условия, то предполагается, что в период логарифмической фазы роста в течение бесконечно малого промежутка времени dt увеличение биомассы dх должно быть пропорционально количеству биомассы х и интервалу времени, то есть
dх = μх dt, (1)
откуда
dх/dt = μх (2)
Дифференциальное отношение dх/dt выражает скорость роста популяции. Параметр μ, обозначающий скорость роста единицы биомассы (1/х)(dх/dt), называется удельной скоростью роста и измеряется в единицах, обратных времени (1/t). Этот параметр аналогичен сложным процентам. Так, например, удельная скорость роста 0,1 ч-1 эквивалентна скорости 10 % в час.
Если μ постоянна, то интегрирование уравнения (2) дает
1n х = 1n х0 + μt, (3)
где х0 - биомасса в начальный момент времени t=0. Г рафик зависимости 1n х от времени будет иметь вид прямой линии с наклоном μ.
Из уравнения (3) следует, что
1n (х/ х0) = μt (4)
тогда
μ = 1n (х/х0): t, (5)
а
х = х0 е μt (6)
Рост, подчиняющийся этому закону, называется экспоненциальным, или логарифмическим ростом. Основным параметром, характеризующим скорость роста, служит удельная скорость роста. Остальные параметры роста во многих случаях могут быть выражены через удельную скорость роста.
5.2.2. Время удвоения биомассы
Зависимость между удельной скоростью роста и временем удвоения (td) биомассы можно получить, подставив в уравнение (4)
х = 2 х0 и t = td,
тогда
td = ln2 : μ = 0,693 : μ (7)
5.2.3. Степень размножения
Степень размножения определяется отношением х/х0, которое равно еμt. Если биомасса претерпевает п удвоений или генераций, то можно записать:
х/х0 = 2n (8)
Таким образом,
n = 3,32 log(x/x0) (9)
5.2.4. Экономический коэффициент
Если рост микробной популяции ограничен определенным субстратом, то между начальной концентрацией внесенного в среду лимитирующего субстрата и полученным общим урожаем существует линейная зависимость. Поэтому масса клеток, образованная на единицу лимитирующего субстрата, представляет собой константу - экономический коэффициент, или выход биомассы (Y). Величина Y может быть определена из уравнения:
∆х/∆у = Y, (10)
где ∆х - увеличение биомассы, соответствующее потреблению субстрата в количестве ∆у. Экономический коэффициент, определяется пределом, к которому стремится отношение ∆х/∆у при ∆s —> 0, то есть
Y = dх/ ds (11)
Если х и s - концентрация биомассы и субстрата соответственно, то более строго:
Y = - dх/ ds.
Знак минус вводится потому, что значения х и s изменяются в разные стороны.
Если внешние условия в бактериальной культуре поддерживаются постоянными, то экономический коэффициент тоже будет постоянной, количественно воспроизводимой величиной. Таким образом, если х0 и s0 - начальные концентрации биомассы и субстрата соответственно, а х и s - соответствующие концентрации во время роста в культуре, то
х - х0 = Y (s0 - s) (12)
отсюда
Y = (х - Х0) / (s0 - s) (13)
Экономический коэффициент, измеренный исходя из потребленного органического субстрата, становится показателем эффективности превращения субстрата в бактериальную массу.
5.2.5. Метаболический коэффициент
Скорость потребления субстрата культурой в данный момент времени выражается отношением:
ds/dt = qх, (14)
где х - биомасса, а коэффициент q известен как метаболический коэффициент или удельная скорость метаболизма. Метаболический коэффициент аналогичен ферментативной активности. Если состав биомассы и окружающая среда постоянны, то и д должен быть величиной постоянной.
Для потребления субстрата в небольшой промежуток времени dt можно записать
ds = (μ х : Y) dt, (15)
откуда
ds/dt = μ х/Y (16)
Сравнивая уравнения (16) и (14), становится видно, что
q =μ /Y (17)
Уравнение (17) используется для определения потребностей в субстрате, особенно в кислороде, при различных скоростях роста.
5.2.6. Урожай биомассы
Под урожаем биомассы понимают разность между максимальной и исходной массой бактерий: Х = Хмакс - Х0. Эту величину выражают в граммах сухого вещества.