Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Мембранные структуры клетки
Жидкостно-мозаичная модель строения биомембран
Понимание того, что плазматическая мембрана представляет собой вполне определённую структуру, было достигнуто в середине XIX столетия. В конце XIX столетия Овертон обратил внимание на корреляцию между скоростью, с которой небольшие молекулы проникают в растительные клетки, и их коэффициентом распределения между маслом и водой; это привело его к мысли о липидной природе мембран.
В 1925 г. Гортер и Грендел предположили, что липиды в мембране эритроцитов образуют бимолекулярный слой (липидный бислой) (рисунок 3(a)).
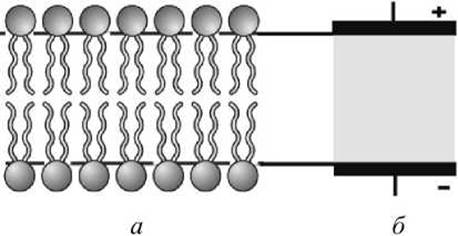
Рисунок 3 - Конденсаторная модель липидного бислоя: а - бимолекулярный слой липидов, б - мембрана как конденсатор
Модель Гортера и Грендела возникла на основе результатов простого эксперимента. Липиды эритроцитов экстрагировали ацетоном и затем в кювете Лэнгмюра получали из них тонкую пленку на поверхности воды. С помощью поплавка сжимали слой липидных молекул на границе раздела вода-воздух до тех пор, пока этот слой не начинал оказывать
сопротивление дальнейшему сжатию; это явление было объяснено образованием плотноупакованной мономолекулярной липидной пленки.
Измерение площади, занимаемой липидами, и сравнение её с площадью поверхности эритроцитов, из которых эти липиды были экстрагированы, дали соотношение 2:1. Отсюда был сделан вывод, что мембрана эритроцитов состоит из липидных молекул, расположенных в два слоя. В историческом плане эта работа имела большое значение, поскольку с тех пор концепция липидного бислоя как структурной основы биологических мембран стала доминирующей и на самом деле оказалась верной. Эту гипотезу подтвердили в 1935 г. Коул и Кертис определив электрические параметры биологических мембран: высокое электрическое сопротивление R ≈ 107 Oм∙м2 и большую удельную ёмкость Cs≈ 0,5 ∙ 10-2 Ф/м2.
Биологическую мембрану можно рассматривать как электрический конденсатор (рисунок 3(6)), в котором пластинами являются электролиты наружного и внутреннего растворов (внеклеточного и цитоплазмы) с погружёнными в них головами липидных молекул. Проводники разделены диэлектрическим слоем, образованным неполярной частью липидных молекул - двойным слоем их хвостов. Диэлектрическая проницаемость липидного слоя s = 2. Ёмкость плоского конденсатора
![]()
где ε0 = 8,85 ∙ 1012 Ф/м - электрическая постоянная; d - расстояние между пластинами конденсатора; S - площадь пластины.
Удельная ёмкость (ёмкость, приходящаяся на единицу площади) определяется как
![]()
Поэтому можно рассчитать расстояние между пластинами конденсатора, соответствующее в нашем случае толщине липидной части мембраны
![]()
Это по порядку величины соответствует наблюдаемой толщине мембран.
Кроме липидов в состав мембраны входят белковые молекулы. Так, коэффициент поверхностного натяжения клеточных мембран значительно ближе к коэффициенту поверхностного натяжения на границе раздела белок-вода (~10-4 Н/м), чем на границе раздела липид-вода (~10-2 Н/м).
Концепция бислойной липидной мембраны получила дальнейшее развитие в предложенной в 1935 г. модели Дэвсона-Даниелли, или модели "сэндвича" ("бутербродная" модель). Согласно этой модели мембрана состоит из трёх слоёв: два крайних слоя состоят из белковых молекул, а средний слой - липидный.
Сингер и Никольсон в 1972 г. на основе совокупности результатов, полученных физическими и химическими методами исследования такими, как рентгеноструктурный анализ и электронная микроскопия, предложили жидкостно-мозаичную модель строения биологических мембран, которая является общепринятой в настоящее время (рисунок 4).
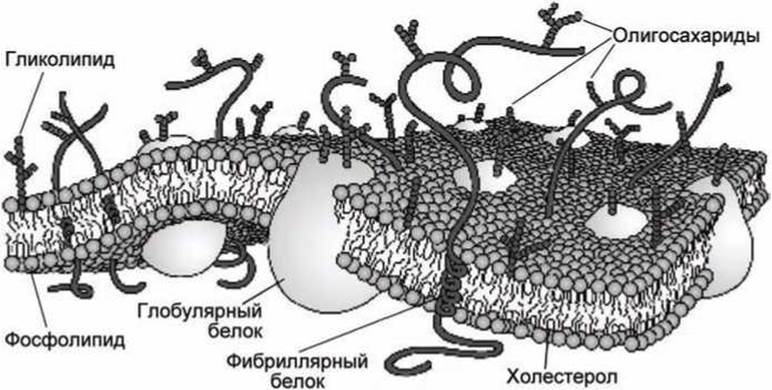
Рисунок 4 - Жидкостно-мозаичная модель плазматической мембраны
В этой модели структурную основу биологической мембраны образует двойной слой фосфолипидов, в котором располагаются белковые молекулы. Липиды находятся при физиологических условиях в жидком агрегатном состоянии. Это позволяет сравнить мембрану с фосфолипидным морем, по которому плавают белковые "айсберги".
Фосфолипиды клетки формируют липидные бислои самопроизвольно, в результате процесса фазового разделения, при котором понижается свободная энергия раствора фосфолипидов в воде. Процесс самопроизвольного формирования биомембран называют самосборкой.
Углеводородные хвосты фосфолипидов обоих слоёв бислоя образуют гидрофобную внутреннюю часть биомембраны толщиной 3-4 нм - гидрофобный углеводородный слой (рисунок 5).
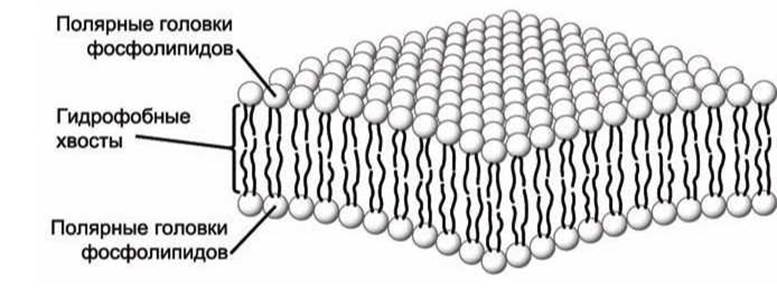
Рисунок 5 - Схема фосфолипидного бислоя
На электронных микрофотографиях сечение мембраны окрашенной молекулами тетрооксида осьмия (которые присоединяются к полярным головкам фосфолипидов) выглядит как "железнодорожное полотно", два "рельса" которого соответствуют двум монослоям бислоя (рисунок 6).
Липидный бислой обладает двумя важными свойствами.
1. Гидрофобный углеводородный слой является непреодолимым барьером, через который невозможна самопроизвольная диффузия водорастворимых (гидрофильных) молекул и ионов. Важно, что этим барьерным свойством можно управлять с помощью мембранных белков, которые осуществляют транспорт (перенос, транслокацию) через мембрану тех веществ, для самопроизвольной диффузии которых биомембрана непроницаема.

Рисунок 6 - Электронная микрофотография среза клетки вблизи плазматической мембраны
2. Биомембрана является стабильным образованием. Бислойность биомембраны определяется гидрофобными и ван-дер-ваальсовыми взаимодействиями между липидами. И даже если состояние внешней среды (например, ионная сила или pH) изменится значительно, бислой имеет достаточный "запас прочности" для сохранения цельности и непроницаемости.
Природные мембраны различных клеток демонстрируют огромное разнообразие, которое определяется функциями клеток данного типа (рисунки 7, 8).
Гладкая гибкая поверхность плазматической мембраны эритроцитов позволяет этим клеткам "протискиваться" через капилляры кровеносной системы, диаметр которых вдвое меньше диаметра дискообразной клетки эритроцита.
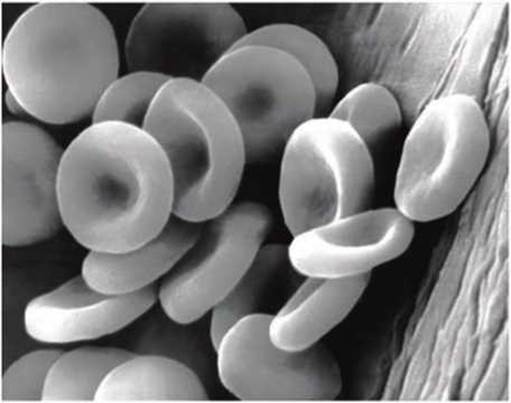
Рисунок 7 - Эритроциты имеют гладкую наружную мембрану
Некоторые клетки имеют длинные тонкие "отростки" плазматической мембраны - реснички и жгутики, которые совершают периодические волнообразные движения (рисунок 8).

Рисунок 8 - Пучки ресничек на поверхности клеток эпендимного эпителия головного мозга защищают их от повреждений
Такое движение стимулирует течение окружающей жидкости по поверхности эпителия. Или, наоборот, как в случае спермиев, способствуют движению самих клеток в окружающей среде.
Аксоны многих нейронов "упакованы" в оболочку из множества слоёв модифицированной плазматической мембраны, которая называется миелиновой оболочкой (рисунок 9). Эти мембранообразные структуры вырабатываются окружающими глиальными клетками и обеспечивают передачу нервных импульсов на большие расстояния.
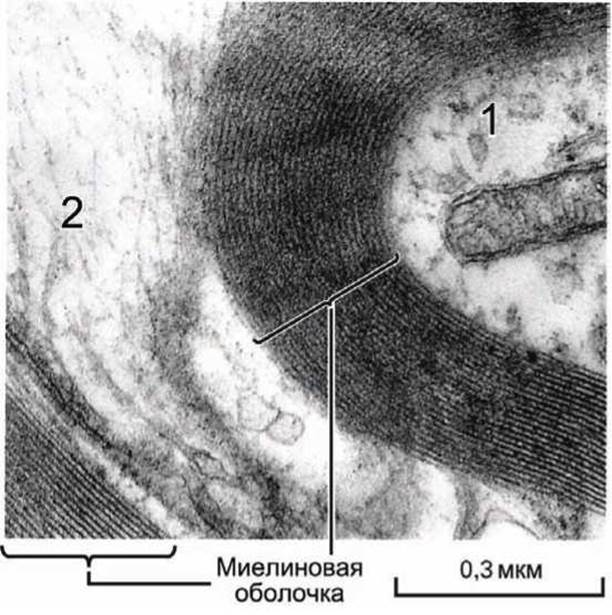
Рисунок 9 - Миелиновая оболочка аксонов (1) позвоночных формируется окружающими опорными глиальными (2) клетками
Несмотря на то, что в приведённых примерах биомембраны имеют различную форму и функции, структурно и эти, и все другие биомембраны устроены одинаково - все они являются липидными бислоями.
Поскольку все клеточные мембраны полностью окружают всю клетку или весь внутренний компартмент, то у всех биомембран одна из поверхностей всегда ориентирована внутрь компартмента - она называется внутренняя поверхность (или внутренний слой бислоя), а другая поверхность всегда экспонирована наружу - она называется наружная поверхность (или наружный слой бислоя). Часто также употребляют названия цитозольная сторона и экзоплазматическая сторона биомембраны, соответственно.
Такие названия удобны для обозначения топологической эквивалентности различных сторон различных мембран (рисунок 10).
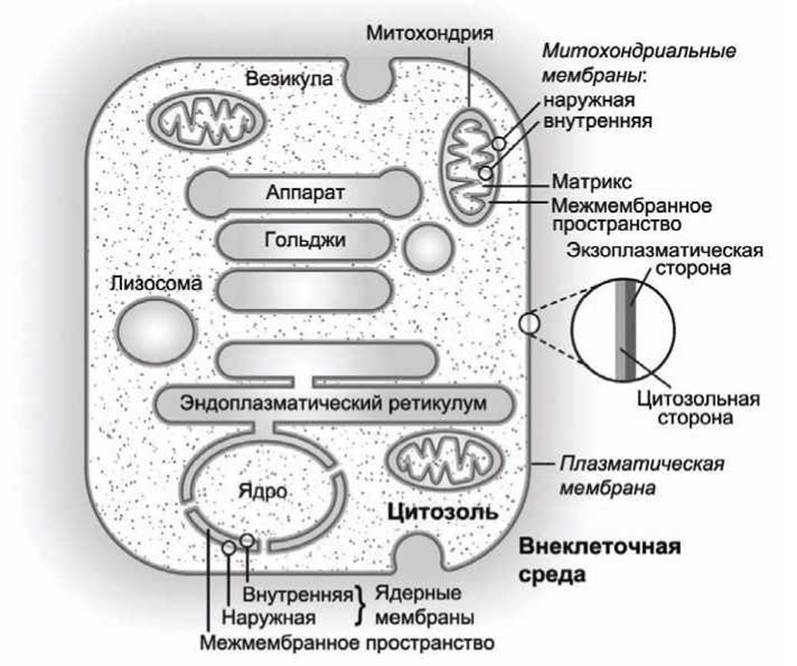
Рисунок 10 - Ориентация цитозольной и экзоплазматической сторон различных мембран клетки
Например, экзоплазматическая сторона плазматической мембраны ориентирована наружу от цитоплазмы в сторону внеклеточного пространства, и она является внешней границей клетки.
Но, в отличие от плазматической мембраны, для органелл и везикул, имеющих только одну мембрану, экзоплазматическая сторона этой мембраны ориентирована внутрь органеллы, она находится в контакте с внутренним содержимым органеллы, которое в этом случае, по отношению к цитозолю клетки, эквивалентно внеклеточной жидкости.
Эта эквивалентность наиболее наглядно проявляется в случае эндоцитозных везикул, которые образуются при "впячивании" участка плазматической мембраны, в результате чего внешняя сторона плазматической мембраны становится внутренней стороной мембраны везикулы.
Различие цитозольной и экзоплазматической сторон биомембраны имеет важное функциональное значение, поскольку при любых перемещениях в составе мембраны цитозольные домены мембранных белков всегда остаются в цитозоле, сохраняя тем самым заданную ориентацию белка относительно компартментов клетки.
Три типа клеточных органелл - ядро клетки, митохондрии и хлоропласты - окружены двумя мембранами. В этих органеллах экзоплазматическая сторона мембран ориентирована в сторону межмембранного пространства.
Модель строения биомембраны Сингера и Никольсона (жидкостномозаичная) в настоящее время общепринята. Однако, как всякая модель, она даёт довольно упрощённую картину строения мембраны. В частности, обнаружено, что белковые "айсберги" не всегда свободно "плавают" в липидном море, а могут быть "заякорены" на внутренние (цитоплазматические) структуры клетки. Кроме того, не все липиды в мембране расположены по принципу бислоя. Липидная фаза мембран содержит также участки, в которых липидные молекулы не образуют двойной слой. Выявлен также специфический класс амфифильных белков, которые под влиянием внеклеточных сигналов меняют свою гидрофобность и обратимо диссоциируют от мембраны.
Таким образом, клеточная мембрана всё более отличается по своим свойствам от "классического" липидного бислоя. Тем не менее, жидкостно-мозаичная модель строения биомембраны Сингера и Никольсона в её разных модификациях всё ещё служит в качестве концептуальной основы для объяснения многих мембранных феноменов. Сложность создания единой модели биологических мембран связана с огромным разнообразием мембранных функций.