МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 6. ОСОБЛИВОСТІ ФІЗІОЛОГІЇ АНАЕРОБНИХ МІКРООРГАНІЗМІВ
Бродіння
Бродіння - це спосіб отримання енергії, при якому АТФ утворюється у процесах анаеробного окиснення органічних субстратів у результаті субстратного фосфорилювання без участі неорганічних акцепторів електронів. Донори та акцептори електронів при бродінні формуються з органічних субстратів із середнім ступенем окиснення, таких як цукри, органічні кислоти, амінокислоти та гетероциклічні сполуки, у процесі їхнього збродження до СО2, молекулярного водню, форміату, ацетату та інших коротколанцюгових жирних кислот. Це найдавніший і найпримітивніший спосіб отримання енергії. Він притаманний деяким групам анаеробних і факультативно анаеробних мікроорганізмів, а саме: молочнокислим, пропіоновокислим бактеріям, клостридіям, ентеробактеріям, дріжджам та ін. Примітивність процесів бродіння полягає в тому, що при розщепленні субстрату вивільнюється лише незначна кількість хімічної енергії, а більшість її залишається в кінцевих продуктах. Бактерії-бродильники отримують енергію лише за рахунок субстратного фосфорилювання. Цикл Кребса в них не бере участь у процесах катаболізму, а дихальний ланцюг не функціонує або зовсім відсутній. Тому при розщепленні однієї молекули глюкози, наприклад при гомоферментативному молочнокислому бродінні, синтезується лише дві молекули АТФ (для порівняння: при аеробному диханні на 1 молекулу глюкози утворюється 38 АТФ).
Незважаючи на те, що зброджуватися може велика кількість субстратів, кількість реакцій, які призводять до безпосереднього синтезу АТФ під час бродіння, досить обмежена. Найпоширеніші з них наведено на рис 6.1.
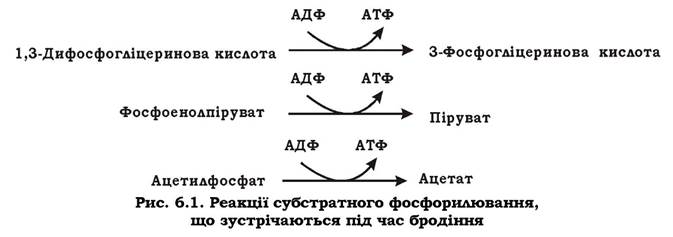
Термінальними акцепторами водню у процесі бродіння, як правило, є органічні метаболіти, що утворилися в результаті розщеплення субстрату. Відновлені сполуки виділяються з клітини і накопичуються в середовищі у значних кількостях. Через низький енергетичний вихід процесів бродіння для забезпечення енергією всіх функцій та біосинтетичних процесів клітині доводиться переробляти величезну кількість субстрату.
Молочнокисле бродіння. У результаті молочнокислого збродження вуглеводів утворюється молочна кислота. Цей процес здійснюють молочнокислі бактерії. Розрізняють гомо- й гетероферментативне молочнокислі бродіння. При гомоферментативному молочнокислому бродінні основним кінцевим продуктом є лактат. Цей процес здійснюють такі бактерії, як Lactococcus lactis, Pediococcus cerevisiae, Lactobacillus acidophilus, Lactobacillus bulgancus. При гетероферментативному молочнокислому бродінні, окрім лактату, утворюється ряд інших кінцевих продуктів: етанол, ацетат, СО2, маніт, причому спектр і кількість метаболітів залежить від виду бактерій та субстрату, що зброджується. Гетероферментативне молочнокисле бродіння притаманне Lactobacillus plantarum, Lactobacillus brevis, Leuconostoc mesenteroides. У Bifidobacterium bifidum гетероферментативне бродіння відбувається за особливою схемою.
Гомоферментативне молочнокисле бродіння,. Основними субстратами для нього є моносахариди (у першу чергу глюкоза), дисахариди (мальтоза, лактоза) та полісахариди (глікоген, крохмаль).
Перш ніж включитися у процес бродіння, дисахариди перетворюються у фосфорильовані моносахариди за загальною схемою:
а) Лактоза → D-Глюкоза + D-Галактоза
D-Галактоза + АТФ → D-Галактозо-1-фосфат + АДФ
D-Галактозо-1-фосфат → Глюкозо-1-фосфат
У цьому випадку глюкозо-1-фосфат утворюється з витратою АТФ.
б) Мальтоза + Н3РО4 → Глюкозо-1-фосфат + Глюкоза
У цьому випадку глюкозо-1-фосфат утворюється без витрат АТФ.
Полісахариди попередньо гідролізуються, а глюкозний залишок фосфорилюється до глюкозо-1-фосфату. Глюкозо-1-фосфат, що утворився при підготовчих перетвореннях вуглеводів, згодом перетворюється у глюкозо-6-фосфат. Подальше перетворення глюкозо-6-фосфату відбувається за єдиною схемою, незалежно від його походження (рис. 6.2).
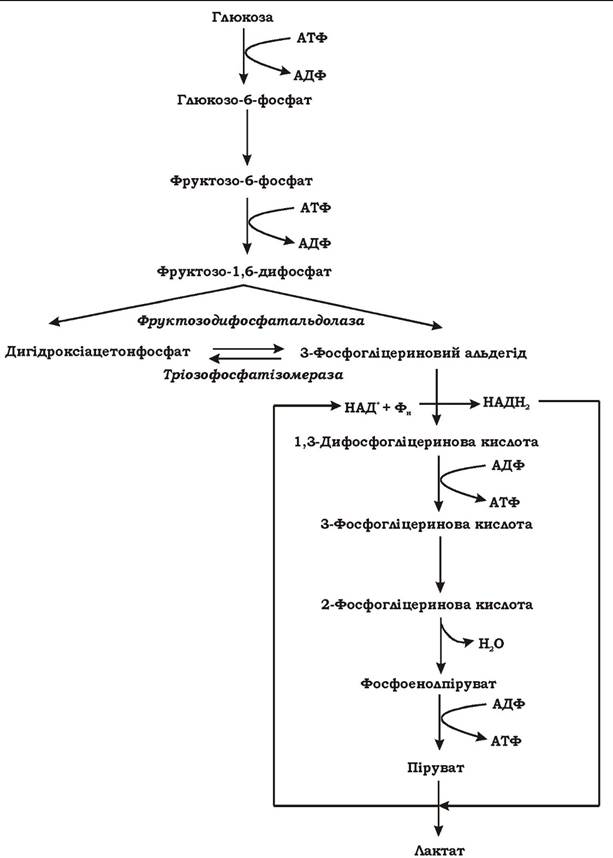
Рис. 6.2. Загальна схема гомоферментативного молочнокислого бродіння
Глюкозо-6-фосфат окиснюється до пірувату гліколітичним шляхом. При цьому утворюється чотири молекули АТФ. Оскільки у процесі гліколізу 2 АТФ витрачається на фосфорилювання глюкози та фруктозо- 6-фосфату, загальний енергетичний вихід процесу становить 2 АТФ на одну молекулу глюкози. Термінальним акцептором електронів при цьому типі бродіння є піруват. Відновлюючись за рахунок НАДН2, піруват перетворюється в лактат, який виділяється з клітини в середовище. При гомофермертативному молочнокислому бродінні лактат є практично єдиним кінцевим метаболітом.
Гетероферментативне (фосфокетолазне) молочнокисле бродіння. У бактерій, що здійснюють цей тип бродіння, відсутні такі ферменти гліколітичного шляху, як фруктозодифосфатальдолаза та тріозофосфатізомераза (рис. 6.3), тому спочатку збродження глюкози в них відбувається за пентозофосфатним шляхом (рис. 6.4). За рахунок пентозофосфатного шляху виділяється СО2, відновлюється дві молекули НАД+ (НАДФ+) та утворюється ксилулозо-5-фосфат. Наступним етапом є розщеплення ксилулозо-5-фосфату на 3-фосфогліцериновий альдегід та ацетилфосфат за допомогою ключового ферменту цього шляху - тіаміндифосфат-залежної фосфокетолази.
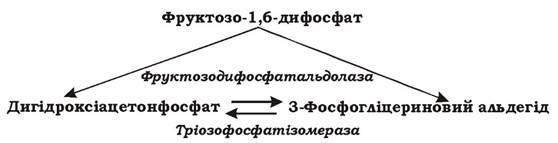
Рис. 6.3. Ферменти, що відсутні в гетероферментативних молочнокислих бактерій
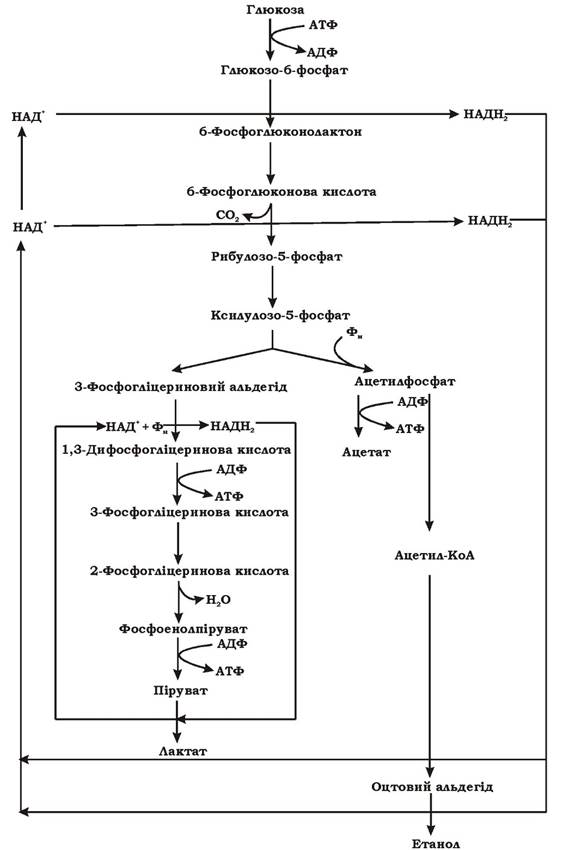
Рис. 6.4. Схема гетероферментативного молочнокислого бродіння, що спричиняється біфідобактеріями
3-фосфогліцериновий альдегід у подальшому окиснюється за допомогою ферментів гліколітичного шляху до пірувату (з утворенням 2 АТФ за рахунок субстратного фосфорилювання), а піруват служить акцептором водню і відновлюється до молочної кислоти. Тобто ця частина гетероферментативного молочнокислого бродіння є фрагментом гомоферментативного молочнокислого бродіння.
Ацетилфосфат перетворюється в ацетат за допомогою ацетаткінази з утворенням АТФ. Крім того, ацетилфосфат може утворювати ацетил-КоА в реакції, що каталізується фосфотрансацетилазою, а згодом відновлюється до оцтового альдегіду та етанолу.
Особливий шлях гетероферментативного молочнокислого бродіння виявлено в Bifidobacterium bifidum (рис. 6.5). У цих бактерій розщеплення та окиснення глюкозо-6-фосфату призводить до утворення пірувату й ацетилфосфату (2С6 → 2С3 + 3С2). Діоксид вуглецю в реакціях даного шляху не утворюється. Спочатку відбувається ізомеризація двох молекул глюкозо-6-фосфату у дві молекули фруктозо-6-фосфату. Потім одна молекула фруктозо-6-фосфату за участю неорганічного фосфату розщеплюється на ацетилфосфат та еритрозо-4-фосфат, а інша вступає в реакцію з еритрозо-4-фосфатом. Продуктами цієї реакції є 3-фосфогліцериновий альдегід та седогептулозо-7-фосфат (С + С4 → С3 + С7). На наступних етапах обидва продукти обмінюються С2-фрагментами, перетворюючись у ксилулозо-5-фосфат та рибозо-5- фосфат (С3 + С7 → 2С5). Рибозо-5-фосфат перетворюється у ксилулозо-5-фосфат. На завершення дві молекули ксилулозо-5-фосфату за участю неорганічного фосфату розщеплюються на дві молекули ацетилфосфату та дві молекули 3-фосфогліцеринового альдегіду. Останні окиснюються до двох молекул пірувату. Піруват відновлюється до лактату, а ацетилфосфат перетворюється в ацетат з виділенням АТФ.
Сумарне рівняння процесу: 2 Глюкоза → 2 Лактат + 3 Ацетат
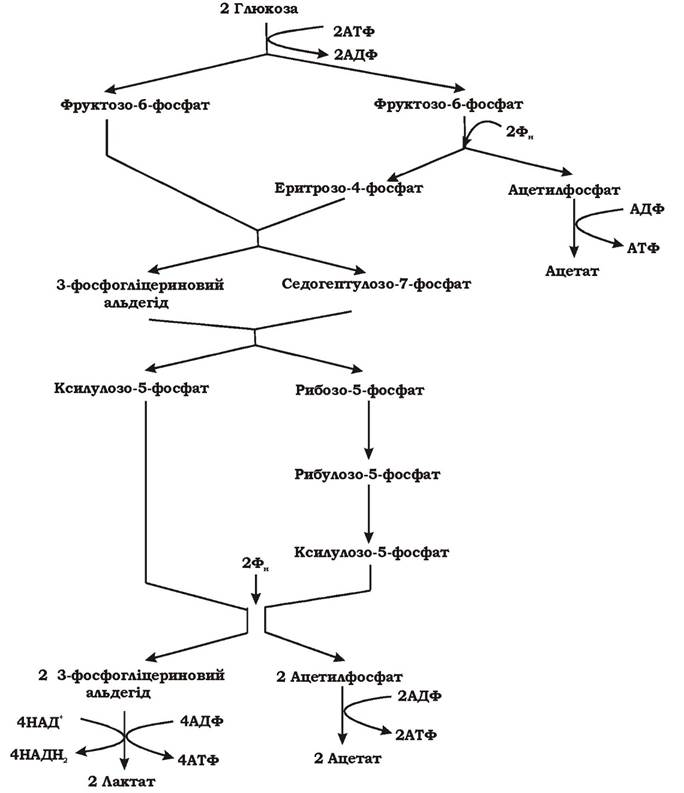
Рис. 6.5. Схема гетероферментативного молочнокислого бродіння, що спричиняється біфідобактеріями
Отже, гетероферментативне молочнокисле бродіння - це анаеробний процес, у результаті якого з глюкози утворюється такі продукти, як молочна, оцтова кислоти, етиловий спирт та вуглекислий газ.
Спиртове (етанольне) бродіння - це анаеробний процес, основним продуктом якого є етиловий спирт. Спиртове бродіння характерне для дріжджів і деяких бактерій, зокрема Sarcina ventriculi, Erwinia amylovora, Zymomonas mobilis. Дріжджі окиснюють глюкозу гліколітичним шляхом, а Zymomonas - шляхом Ентнера - Дудорова.
Дріжджі здійснюють три форми спиртового бродіння за К. Нейбергом:
1. Перша форма - класичне спиртове бродіння, яке відбувається в кислому середовищі (рН 3-6). Воно має два періоди: індукційний і стаціонарний. Оскільки основними продуктами індукційного періоду є гліцерин та піруват, його називають гліцеропіровиноградним бродінням.
2. Друга форма - спиртове бродіння, яке проходить за наявності гідросульфіту натрію. Оскільки основним кінцевим продуктом цього процесу є гліцерин, його ще називають гліцериновим бродінням.
3. Третя форма - спиртове бродіння, що відбувається в лужному середовищі.
Перша формна спиртового бродіння за Нейбергом у загальному вигляді відбувається за схемою, наведеною на рис. 6.6.
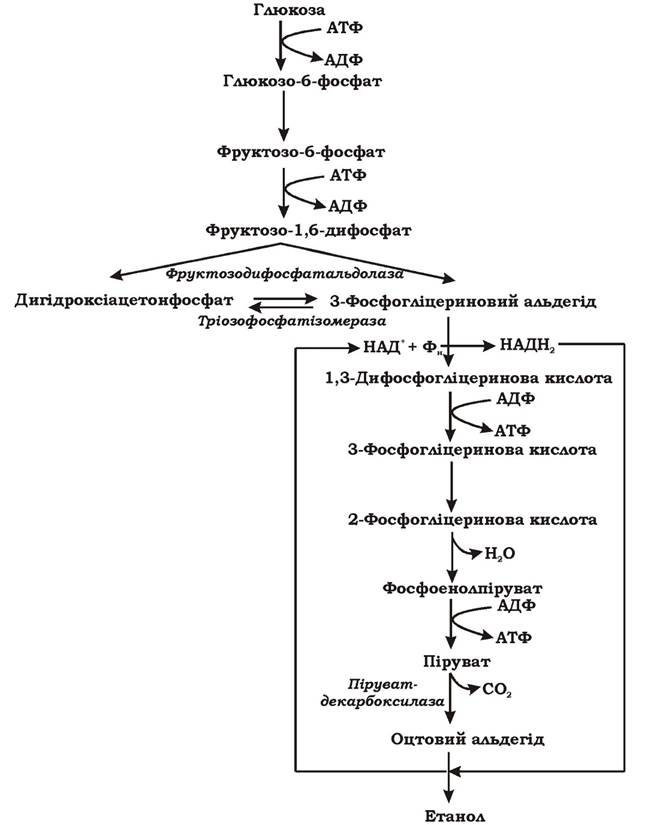
Рис. 6.6. Загальна схема спиртового бродіння (перша форма спиртового бродіння за Нейбергом)
Процес спиртового та гомоферментативного молочнокислого бродіння в цілому подібні. Спочатку глюкоза окиснюється до пірувату гліколітичним шляхом, але на наступних етапах спиртове бродіння має дві відмінності. Перша полягає в тому, що акцептором водню є не безпосередньо піруват, а продукт його декарбоксилювання - оцтовий альдегід. Декарбоксилювання пірувату каталізується ключовим ферментом спиртового бродіння - піруватдекарбоксилазою. Особливістю цієї реакції є її повна незворотність. Оцтовий альдегід відновлюється до етанолу за участю НАД+-залежної алкогольдегідрогенази. Друга відмінність полягає в тому, що при спиртовому бродінні відбувається чітке міжмолекулярне розмежування на відновлені молекули (спирт) і окиснені (СО2), а при гомоферментативному молочнокислому бродінні кінцевий продукт - молочна кислота за сумарним ступенем окисненості-відновленості не відрізняється від молекули гексози.
Як уже зазначалося, спиртове бродіння має два періоди: індукційний та стаціонарний. Індукційний (початковий) період характеризується накопиченням у середовищі гліцерину й пірувату, тому його можна розглядати як окремий вид бродіння - гліцеропіровиноградне.
Висока концентрація глюкози в середовищі інгібує алкогольдегідрогеназу, а кількість оцтового альдегіду на початку спиртового бродіння ще недостатня для індукції синтезу цього ферменту. Тому НАДН2 на цьому етапі переносить водень не на оцтовий альдегід, а на другу молекулу 3-фосфогліцеринового альдегіду. У результаті перша молекула 3-фосфогліцеринового альдегіду окиснюється до пірувату, а друга відновлюється до гліцерину (рис. 6.7).
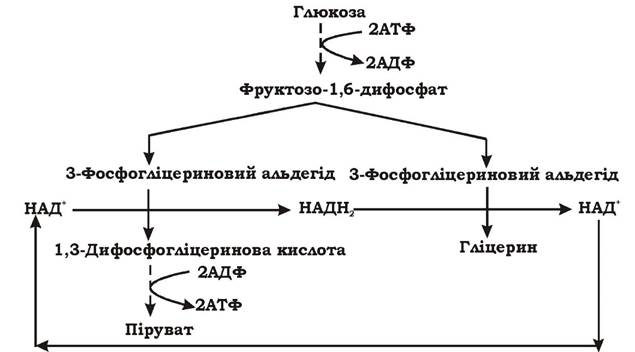
Рис. 6.7. Схема гліцеропіровиноградного бродіння (індукційний період спиртового бродіння)
Стаціонарний період спиртового бродіння починається після зменшення концентрації глюкози в середовищі та достатнього накопичення оцтового альдегіду. Активність алкогольдегідрогенази зростає, і оцтовий альдегід починає відновлюватися воднем до етанолу. Енергетичний вихід процесу - 2 моля АТФ на одну молекулу глюкози.
Друга форма спиртового бродіння за Нейбергом - це бродіння за наявності гідросульфіту натрію. Оскільки основним кінцевим продуктом цього процесу є гліцерин, його ще називають гліцериновим бродінням. Енергетичний вихід гліцеринового бродіння вдвічі менший, ніж класичного спиртового бродіння, бо одна тріоза не окиснюється, а відновлюється до гліцерину. Це пов'язано з тим, що гідросульфіт натрію утворює комплекс з оцтовим альдегідом, який не може бути акцептором водню (рис. 6.8).
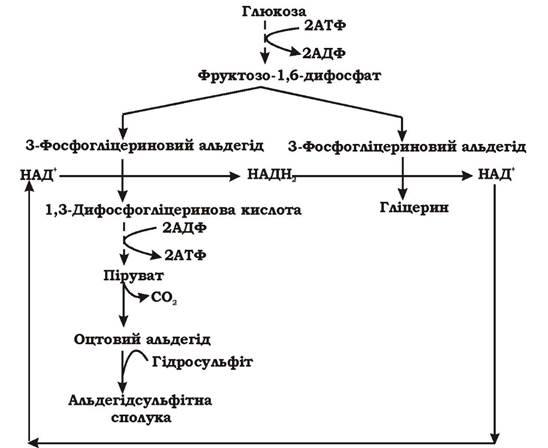
Рис. 6.8. Схема гліцеринового бродіння (друга форма спиртового бродіння за Нейбергом)
Третя форма спиртового бродіння за Нейбергом - це бродіння в лужному середовищі. За цих умов оцтовий альдегід дегідрогеназою не відновлюється, тому акцептором водню є 3-фосфогліцериновий альдегід, як і в разі гліцеропіровиноградного бродіння. Перенесення водню на 3-фосфогліцериновий альдегід приводить до накопичення гліцерину. Крім того, за цих умов дві молекули оцтового альдегіду вступають у реакцію дисмутації: одна молекула окиснюється до оцтової кислоти, а друга відновлюється до етанолу (рис. 6.9).
Загальне рівняння процесу: 2 Глюкоза → 2 Гліцерин + Ацетат + Етанол + 2СО2
Бактерії роду Zymomonas для здійснення спиртового бродіння розщеплюють глюкозу шляхом Ентнера - Дудорова (рис. 6.10). Глюкоза окиснюється до 2-кето-3-дезокси-6-фосфоглюконової кислоти, яка потім розщеплюється до пірувату та 3-фосфогліцеринового альдегіду. 3-фосфогліцериновий альдегід далі окиснюється гліколітичним шляхом до пірувату з утворенням 2 АТФ. Дві молекули пірувату, що утворилися на обох гілках метаболічного перетворення, декарбоксилюються до оцтового альдегіду, який згодом відновлюється до етанолу таким же чином, як і у дріжджів. Загальне рівняння процесу: Глюкоза → 2 Етанол + 2СО2. Енергетичний вихід процесу - одна молекула АТФ на одну молекулу глюкози.
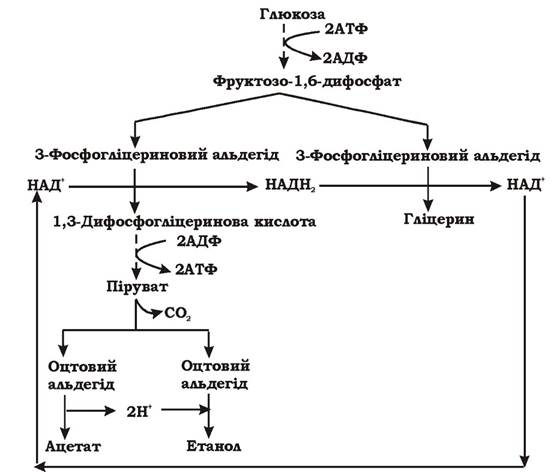
Рис. 6.9. Схема спиртового бродіння в лужному середовищі (третя форма спиртового бродіння за Нейбергом)
Пропіоновокисле бродіння - це анаеробний процес, основним продуктом якого є пропіонова кислота. Крім пропіонату, більшість анаеробів утворюють також ацетат і СО2. Субстратами для пропіоно- вокислого бродіння є глюкоза та лактат. Пропіоновокисле бродіння характерне для бактерій Propionibacterium freudenreichii, P. acidipropionici, P. acnes, Clostridium propionicum, Selenomonas.
У бактерій існує два шляхи утворення пропіонату:
1. Сукцинат-пропіонатний шлях, або шлях метилмалоніл-КоА (характерний для бактерій роду Propionibacterium).
2. Акрилатний шлях (характерний для Clostridium propionicum, Megaspaera elsdenii).
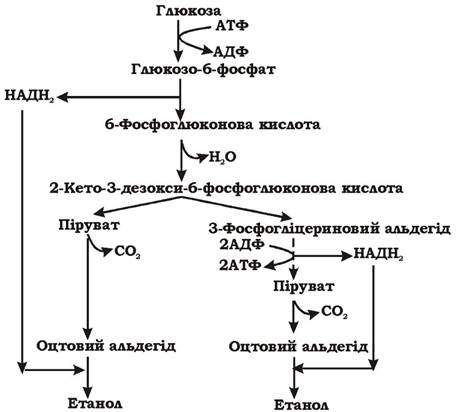
Рис. 6.10. Схема спиртового бродіння, що здійснюють бактерії роду Zymomonas
Схему сукцинат-пропіонатного метаболічного шляху наведено на рис. 6.11. Розшифрувати цей шлях виявилося досить складно, і сьогодні не всі деталі цього процесу повністю зрозумілі. Спочатку глюкоза окиснюється до пірувату гліколітичним шляхом, або лактат окиснюється до пірувату за участю флавопротеїнового переносника водню. Потім піруват перетворюється в щавлево-оцтову кислоту (ЩОК) за реакцією транскарбоксилювання. Донором вуглецю є органічна сполука метилмалоніл-КоА, а переносником СО2 - біотин. Реакція транскарбоксилювання є більш вигідним процесом, ніж пряме карбоксилювання за рахунок неорганічного вуглецю, оскільки здійснюється без витрат енергії. ЩОК служить акцептором водню і відновлюється до фумарату. Подальше відновлення фумарату до сукцинату за допомогою фумаратредуктази пов'язане з синтезом АТФ за участю переносників дихального ланцюга. Цей процес окисного фосфорилювання відомий як фумаратне дихання і характерний для багатьох анаеробних мікроорганізмів. Тобто під час пропіоновокислого бродіння мають місце як виключно бродильні процеси, так і фрагменти анаеробного дихання.
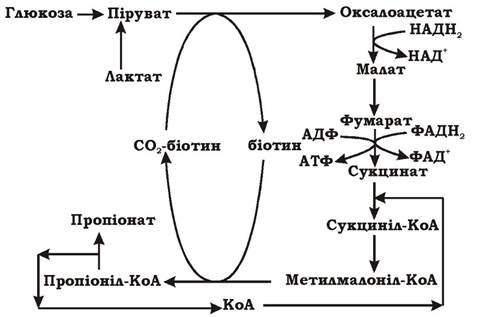
Рис. 6.11. Сукцинат-пропіонатний шлях (шлях метилмалоніл-КоА) утворення пропіонату
У бродінні цього типу беруть участь три КоА-похідні: сукциніл-КоА, метилмалоніл-КоА та пропіоніл-КоА, які взаємно перетворюються за реакціями ізомеризації, що є ключовими у пропіоновокислому бродінні. Ізомерази містять коферменти-Ві2. Отже, для шляху метилмалонілКоА характерні:
- процес циклічного перенесення Сі-фрагмента з метилмалоніл-КоА на піруват з утворенням ЩОК (реакція транскарбоксилювання) без вивільнення СО2 у довкілля;
- процес ізомеризації КоА-похідних і циклічне перенесення КоА з продукту на продукт без вивільнення в середовище;
- фрагменти циклу Кребса, реакції яких здійснюються у зворотному напрямку (не окиснення, а відновлення);
- отримання АТФ субстратним фосфорилюванням під час окиснення глюкози гліколізом та окисним фосфорилюванням у результаті фумаратного дихання.
Акрилатний шлях утворення пропіонату наведено на рис 6.12. Відновлення лактату до пропіонату через акрилатний шлях відбувається спряжено з окисненням лактату до ацетату та СО2. Ключовою реакцією цього шляху є дегідратація лактил-КоА до акриліл-КоА.
Пропіоновокислі бактерії містять гемовмісні ферменти, такі як цитохроми та каталаза. Вони можуть рости і за аеробних умов, але при низьких концентраціях кисню, тому їх відносять до мікроаеротолерантних бактерій.
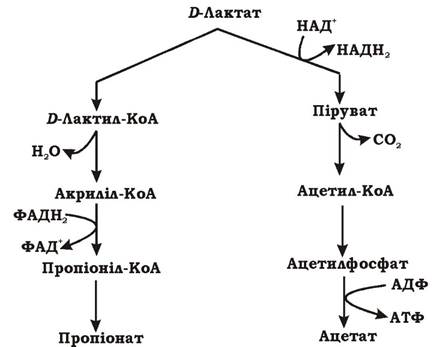
Рис. 6.12. Акрилатний шлях утворення пропіонату
Мурашинокисле (форміатне) бродіння - це анаеробний процес, характерний для ентеробактерій, важливим (але не головним) продуктом якого є мурашина кислота. Крім мурашиної кислоти, утворюється також суміш органічних кислот (молочна, оцтова, бурштинова) та спиртів (етанол, 2,3-бутандіол). Співвідношення кінцевих продуктів залежить від роду бактерій. Бактерії родів Escherichia, Salmonella, Shigella зброджують цукри до молочної, оцтової, бурштинової та мурашиної кислот, СО2, етанолу та Н2. Бактерії родів Enterobacter, Serratia, Erwinia продукують менше кислот, але більше СО2, етанолу і, крім того, великі кількості 2,3-бутандіолу.
Розрізняють два типи мурашинокислого бродіння: змішане (рис. 6.13) та бутандіолове (рис. 6.14).
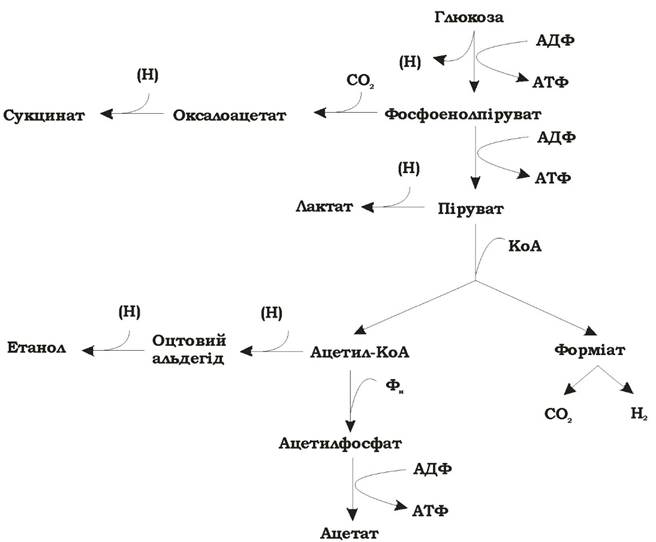
Рис. 6.13. Схема змішаного бродіння
При змішаному та бутандіоловому типах бродіння розщеплення гексози в ентеробактерій іде гліколітичним шляхом. На рівні фосфоенолпірувату (ФЕП) при змішаному бродінні відбувається відгалуження шляху утворення сукцинату; всі решта продуктів утворюється з піру- вату. Піруват метаболізується за участю трьох ферментних систем, і кількість кінцевих продуктів залежить від їхньої активності:
1. Лактат утворюється за допомогою лактатдегідрогенази. Особливо багато лактату накопичується при змішаному бродінні.
2. Перетворення пірувату в ацетил-КоА в ентеробактерій за анаеробних умов здійснює піруватформіатліаза, без відновлення НАД+.
Ацетил-КоА при змішаному та бутандіоловому бродінні в подальшому відновлюється до етанолу, а при змішаному бродінні ще й перетворюється через ацетилфосфат в ацетат з виділенням АТФ.
3. Утворення 2,3-бутандіолу з пірувату при бутандіоловому бродінні відбувається за допомогою а-ацетолактатсинтетази, активність якої зростає при зниженні рН. Так, у Enterobacter aerogenes підкислення середовища до 6,0 сприяє посиленому утворенню 2,3-бутандіолу і зменшенню синтезу органічних кислот.
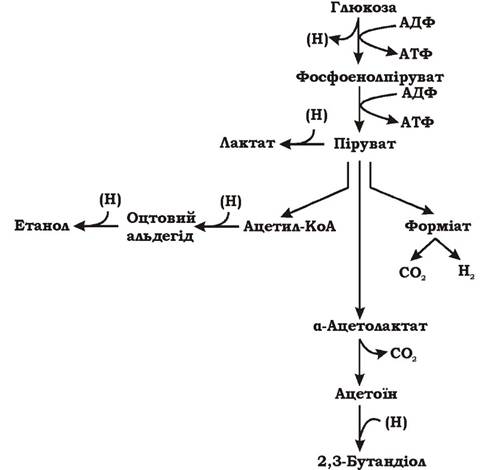
Рис. 6.14. Схема бутандіолового бродіння
Форміат є типовим продуктом мурашинокислого бродіння. Бактерії родів Shigella та Erwinia накопичують значну його кількість у середовищі. Інші представники ентеробактерій, такі як Escherichia coli та Enterobacter aerogenes, містять фермент форміатгідрогенліазу, який розщеплює більшу частину мурашиної кислоти на СО2 та Н2.
Енергетичний вихід мурашинокислого бродіння становить 2-2,5 молекули АТФ на одну молекулу глюкози. Крім 2 молів АТФ, що виділяються при гліколізі, деяка кількість АТФ синтезується при утворенні ацетату з ацетилфосфату за участю ацетаткінази.
Маслянокисле та ацетонобутанолове бродіння - це анаеробний процес перетворення цукрів, унаслідок якого в середовищі можуть накопичуватися масляна кислота (бутират), ацетат, етанол, бутанол, ацетон, ізопропанол, СО2, Н2. Здійснюють його представники роду Clostridium.
На рис. 6.15 наведено загальну схему цього процесу, але шляхи перетворення глюкози і кінцеві продукти залежать від фізіологічних властивостей мікроорганізмів. Маслянокисле та ацетонобутанолове бродіння розглядають як окремі процеси.
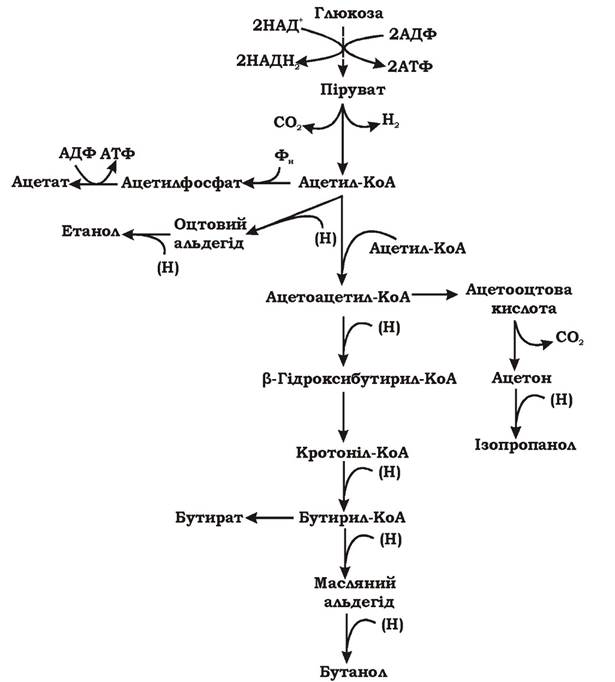
Рис. 6.15. Загальна схема збродження глюкози клостридіями (маслянокисле та ацетонобутанолове бродіння)
Для поглинання цукрів клостридії використовують фосфотрансферазну систему, а розщеплення гексозофосфатів здійснюють гліколітичним шляхом. Важливою реакцією цього бродіння є перетворення пірувату в ацетил-КоА з виділенням молекулярного водню. Цю реакцію називають фосфорокластичною, вона здійснюється в кілька етапів:
1. Спочатку піруват декарбоксилюється за участю ферментної системи піруват:фередоксин-оксидоредуктази, а двовуглецевий фрагмент, що залишився, зв'язується ферментом, який містить тіамінпірофосфат (ТПФ), і перетворюється в гідроксіетилтіамінпірофосфат (ГЕТПФ-Е):
![]()
2. Далі утворюється ацетил-КоА та відновлюється фередоксин:
![]()
3. Фередоксин (низькопотенційний переносник із групи залізосірчаних білків) переносить електрони за допомогою гідрогенази на протони, які походять з води. Саме ці протони виконують роль акцепторів електронів у реакції, в результаті якої виділяється молекулярний водень (рис. 6.16).
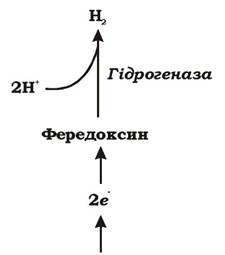
Рис. 6.16. Утворення молекулярного водню при окисненні пірувату
Ацетил-КоА, що утворився при декарбоксилюванні пірувату, є проміжним продуктом при утворенні ацетату та етанолу. Ацетат утворюється з ацетилфосфату, з виділенням АТФ а етанол - у результаті відновлення ацетил-КоА й оцтового альдегіду.
Масляна кислота, бутанол, ацетон і ізопропанол походять з ацетоацетил-КоА, що є продуктом конденсації двох молекул ацетил-КоА за участю тіолази. Ацетоацетил-КоА відновлюється НАДН2 до масляної кислоти та бутанолу. Крім того, ацетоацетил-КоА є попередником утворення ацетону та ізопропанолу.
Механізм збродження амінокислот клостридіями відрізняється від механізму збродження цукрів тим, що донори та акцептори електронів не утворюються з попереднього органічного субстрату. При збродженні амінокислот молекула однієї амінокислоти слугує донором електронів, а молекула іншої - акцептором (реакція Стікланда, рис. 6.17), або дві молекули однієї і тієї ж амінокислоти є донором і акцептором електронів. Кінцевими продуктами збродження амінокислот можуть бути аміак, СО2, молекулярний водень та коротколанцюгові жирні кислоти, які зазвичай зберігають вуглецевий скелет початкової амінокислоти. Енергія при цьому запасається шляхом субстратного фосфорилювання.
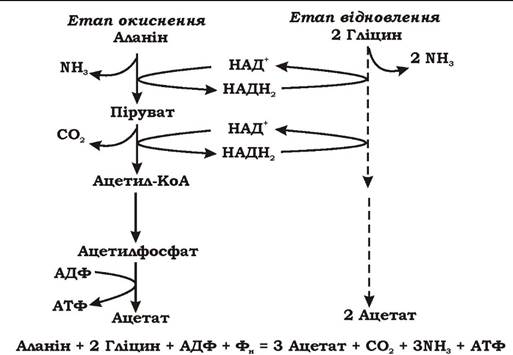
Рис. 6.17. Парне збродження амінокислот за реакцією Стікланда
Процес збродження амінокислот складається з кількох анаеробних окисно-відновних реакцій. Окисний етап включає реакції дезамінування, декарбоксилювання і завжди супроводжується субстратним фосфорилюванням. Відновлення амінокислот - досить складна реакція. Відповідні ферменти зв'язані з мембраною і складаються з кількох білків. На цьому етапі бродіння АТФ практично ніколи не утворюється. Винятком є збродження гліцину, коли субстратне фосфорилювання пов'язане з реакціями відновлення.
Синтрофне (вторинне) бродіння. У результаті розглянутих вище класичних (первинних) бродильних процесів, цукри, органічні кислоти, амінокислоти та гетероциклічні сполуки зброджуються до коротколанцюгових жирних кислот і спиртів. Жирні кислоти, що мають більше двох атомів вуглецю (пропіонат, бутират), спирти (етанол, пропанол, 2,3-бутандіол), а також розгалужені жирні кислоти та ароматичні сполуки є субстратами для вторинних бактерій-бродильників, які ще називають синтрофними. Синтрофні бактерії - облігатні Н+-редуктори, тобто акцепторами електронів для них є протони. Внаслідок відновлення протонів утворюється молекулярний водень, а в результаті окиснення субстрату - ацетат, СО2 і, можливо, фумарат.
Бутират- + 2Н2О → 2 Ацетат-+ Н+ + 2Н2↑ (рис. 6.18)
Пропіонат- + 2Н2О → Ацетат- + СО2 + ЗН2↑ (рис. 6.19)
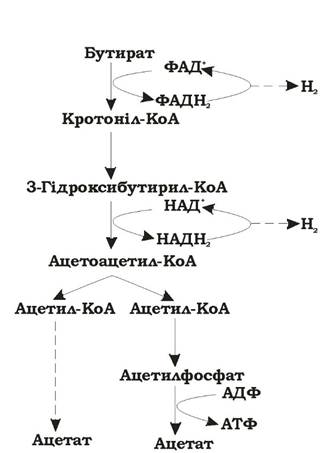
Рис. 6.18. Розщеплення бутирату культурою Syntrophomonas wolfei (молекулярний водень поглинається хемолітотрофами і в середовищі не накопичується)
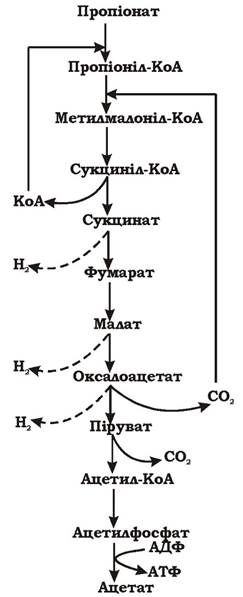
Рис. 6.19. Окиснення пропіонату синтрофними бактеріями (молекулярний водень поглинається хемолітотрофами і в середовищі не накопичується)
Оскільки синтрофні бактерії використовують як донори електронів нижчі жирні кислоти (Ео'> 320 мВ) і як акцептори електронів - протони (Ео'= -420 мВ), ці процеси бродіння є ендергонічними. Однак при низькому тиску Н2 (нижче 1 Па) вторинне бродіння стає енергетично вигідним і забезпечує синтез АТФ. Тому синтрофні бактерії у природі існують лише в синтрофних асоціаціях із літотрофними мікроорганізмами, що поглинають молекулярний водень (метаногенами, хемолі- тотрофними сульфатредукторами, фототрофними зеленими сіркобактеріями, гомоацетогенами). Перенесення молекулярного водню від синтрофних бактерій до бактерій, що його поглинають, можливе лише за умови прямого контакту між клітинами різних видів. При цьому газ у середовище не виділяється. Даний феномен отримав назву міжвидового перенесення водню.
Синтрофія (синтроф - у перекладі з грец. - годувати разом) - це особливий тип симбіотичної взаємодії між двома різними типами бактерій, які з енергетичних причин залежать один від одного при розщепленні певного субстрату.
Представниками синтрофних бактерій є Thermoanaerobium brockii, Pelobacter venetianus, P. acetylenicus, P. carbinolicus (окиснюють етанол, пропанол, ацетилен, 2,3-бутандіол); Syntrophomonas wolfei, S. sapovorans, Syntrophospora bryantii (окиснюють жирні кислоти С4-С11), Syntrophobacter wolinii, S. pfennigii (окиснюють пропіонат, піруват), Clostridiumultunense (окиснюють ацетат, етанол, етиленгліколь, форміат), Syntrophobotulus glycolicus (окиснюють гліколят, гліоксилат), Syntrophus buswellii, S. gentianae (окиснюють бензоат, кротонат).