МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
РОЗДІЛ 7. СИСТЕМАТИКА МІКРООРГАНІЗМІВ
Принципи класифікації бактерій
Перша спроба наукової класифікації мікроорганізмів належить датському зоологу О. Мюллеру, який визначив усі головні форми бактерій. У запропонованій ним класифікації всі відомі на той час види він розділив на два роди: Monas і Vibrio.
Х. Еренберг (1795-1876) у книзі "Інфузорії як досконалі організми" (1838) розділив клас інфузорії на 22 родини, три з яких об'єднували бактерії. У нього зустрічаємо назви Spirillum, Spirochaeta і Bacterium, які вживаються і в сучасній систематиці. Він уперше використав для назв видів мікроорганізмів принцип бінарної номенклатури. З відомих у наш час видів знаходимо в Еренберга опис сінної палички (Bacillus subtilis), Spirillum volutans. Він же вперше описав "чудову паличку" (Bacterium prodigiosum, сучасна назва - Serratia marcescens),
яка викликала жах своєю появою у вигляді кривавих плям на крохмалистих продуктах ("кривавий хліб»).
Перші класифікаційні схеми бактерій будувалися за принципом каталогізації, коли об'єднання мікроорганізмів у таксони здійснювалось на підставі морфологічних ознак. Але з часом стало зрозуміло, що вивчення лише морфологічних характеристик недостатньо для задовільного розподілу бактерій по таксономічних групах. Необхідність використання для класифікації бактерій поряд з морфологічними фізіолого-біохімічних ознак була обґрунтована С. Виноградським і М. Бейєринком.
Вивчення морфолого-культуральних властивостей досліджуваного мікроорганізму - це перший крок у спробі його класифікації. При цьому звертається увага на форму та розміри клітини, морфологію колоній (рис. 7.5), характер росту на рідких поживних середовищах.
Останніми роками в класифікації прокаріотів намітилось два основних напрями. В основу першого покладено ідею створення філогенетичної системи бактерій, тобто побудови єдиної системи, яка б об'єктивно відбивала родинні зв'язки між різними групами прокаріотів з урахуванням їхнього історичного розвитку. Другий напрям передбачає практичні цілі, щоб класифікація прокаріотів служила їх швидкій ідентифікації, тобто встановлення належності організму до певного таксону. Другий напрям чітко простежується у визначниках бактерій Берджі.
Для розробки філогенетичної системи прокаріотів, подібної до такої в рослин і тварин (традиційне еволюційне дерево), спочатку була спроба використати дані про фенотипові ознаки. Усі ознаки, які використовувались, умовно розподілялись за ступенем їхнього значення. У подальшому, виходячи з важливості ознак об'єкти поділяли на таксономічні групи. Отже, неважко помітити, що структура ієрархічної системи визначається порядком, за яким розміщують ознаки, а сам порядок вибирається довільно (суб'єктивно).
Усе це спонукало систематиків шукати інші підходи для встановлення ступеня спорідненості бактерій. З метою виключення елементів суб'єктивізму було запропоновано нумеричну таксономію, основні принципи якої розробив французький ботанік М. Адансон (17271806) ще в 1757 р. В основу нумеричної таксономії покладено принципи, згідно з якими будь-яка ознака при оцінці особини має однакове значення (рівноцінність ознак); при описі досліджуваної культури необхідно використовувати якнайбільше ознак; ступінь подібності є функцією кількості загальних ознак; оцінка й класифікація особин базується на кореляції ознак.
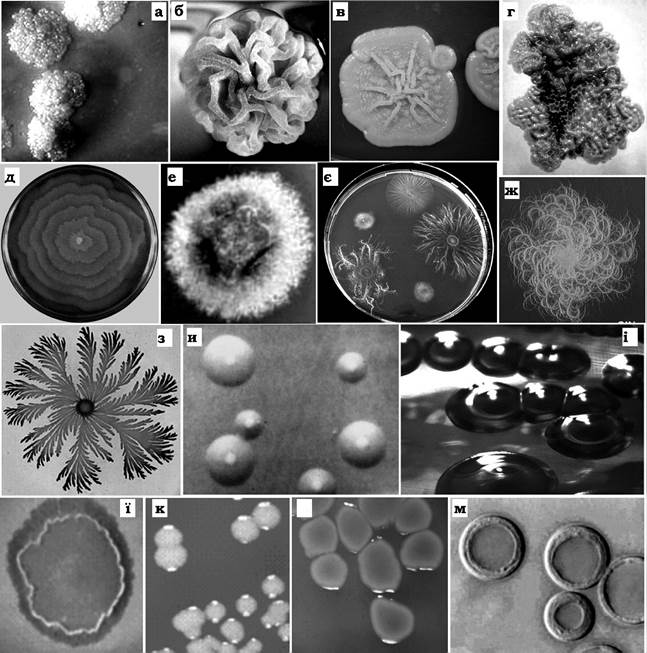
Рис. 7.5. Деякі форми колоній бактерій:
а - пухирчаста; б - складчаста; в - зморшкувата; г - піноподібна; д - концентрична; е - пухнаста; є, ж - нитчаста; з - ризоїдна; и, і - "вилитої яєчні»; ї - пласка із хвилястим валиком по краю; к - кругла, гладенька, блискуча; л - гладенька, неправильної форми; м - кругла з валиком по краю
Класифікація бактерій, побудована за принципами Адансона, досить трудомістка процедура, а тому свій розвиток і практичне використання вона набула з появою ЕОМ. Переваги її полягають у формальній можливості дещо позбутися елементів суб'єктивності, оскільки всі ознаки визнаються рівноцінними. Слабкою стороною цього підходу є
неповна інформативність: для оцінки подібності бактерій використовується близько 100 ознак, що становить приблизно 10 % від тієї кількості, яка визначає бактеріальний фенотип. Цей принцип покладено в основу створення всесвітньо визнаного визначника бактерій Берджі.
Визначники бактерій створюються з урахуванням їхнього практичного використання. За цим принципом підготовлені всі визначники бактерій Д. Берджі (1860-1937). Перше видання вийшло друком у 1923 р., дев'яте видання - "Bergey's Manual of Systematic Bacteriology" - в 1984-1986 рр. Останні видання визначників бактерій готуються Американським товариством бактеріологів із залученням провідних систематиків усього світу. Причому, починаючи з 9-го видання, відбувся поділ цієї попередньо єдиної праці на 4-томну систематику "Bergey's Manual of Systematic Bacteriology" та 2-томний визначник "Bergey's Manual of Determinative Bacteriology".
За дев'ятим виданням, усі бактерії об'єднуються в царство Procaryotae і розділені на чотири основні категорії. Поділ на категорії ґрунтується на наявності і будові клітинної стінки. Кожна категорія об'єднує певні групи бактерій.
Категорія I. Gracilicutes (лат. gracilis - тонкий; cutes - шкіра) - грам- негативні еубактерії, які мають клітинні стінки. Категорія об'єднує бактерії 1-16-ї груп. Клітини можуть бути овальної форми, у вигляді прямих чи зігнутих паличок, спіралей або ниток. Деякі з них можуть бути покриті чохлом або капсулою. Розмноження відбувається переважно шляхом бінарного поділу, інколи - брунькуванням. У деяких представників (пор. Pleurocapsales) розмноження клітин відбувається шляхом множинних внутрішніх поділів з утворенням дрібних сферичних клітин, які називаються беоцитами (наноцитами). Міксобактерії можуть утворювати плодові тіла і міксоспори. Серед представників цього відділу є рухомі форми і такі, що не здатні до руху. Рух може бути "плаваючого" або "ковзного" типу.
Категорія II. Firmicutes (лат. firmus - міцний) - грампозитивні еубактерії, які м.ають клітинні стінки. Категорія об'єднує бактерії 17-29-ї груп. Клітини сферичні, паличко- або ниткоподібні. Інколи палички й нитки галузяться. Розмножуються, як правило, шляхом бінарного поділу. Деякі представники утворюють спори як форми спокою (ендоспори) або спори на гіфах. До цього відділу належать аспорогенні та спороутворювальні бактерії, а також актиноміцети і споріднені з ними організми.
Категорія III. Tenericutes (лат. tener - ніжний, м'який) - еубактерії, позбавлені клітинної стінки. Зазвичай їх називають мікоплазмами і відносять до класу Mollicutes (група 30). Усі вони не здатні до синтезу попередників пептидоглікану. Клітини оточені елементарною мембраною, плеоморфні, їхні розміри варіюють у широких межах. Розмножуються шляхом брунькування, фрагментації, і/або бінарного поділу. Здебільшого це нерухомі форми, але деякі з них проявляють здатність до ковзного руху. За Грамом не забарвлюються. Для свого росту потребують складних середовищ. На щільних поживних середовищах утворюють характерні колонії типу "вилитої яєчні».
Категорія IV. Mendosicutes (лат. mendosus - помилковий) - архебактерії. Прокаріоти, у структурі клітинної стінки яких істинний пептидоглікан не виявляється. Ці бактерії характеризуються різноманітністю метаболізму і здатністю жити в екстремальних умовах. Переважно вони зустрічаються в анаеробних умовах, гіперсолоних або в гідро- і геотермально нагрітих середовищах. Архебактерії можуть бути як мезо-, так і термофілами. Деякі види здатні рости при температурі вище 1000 С. За Грамом забарвлюються позитивно або негативно. У грампозитивних архебактерій клітинна стінка складається з псевдомуреїну, метанохондріотину і гетерополісахариду. Форма клітини може бути сферична, спіральна, пластинчаста або паличкоподібна. Товщина клітини коливається від 0,1 до ~15 мкм, а довжина може досягати 200 мкм.
Розвиток біохімії, молекулярної біології дав змогу розробити інші підходи для встановлення ступеня спорідненості бактерій. Один із них базується на порівняльному вивченні і зіставленні первинної структури макромолекул, які беруть участь у здійсненні найважливіших функцій клітини. До таких макромолекул належать ДНК, РНК, ферментні білки. Відомо, що генетична інформація "записана" в молекулі ДНК у вигляді різних поєднань трьох із чотирьох азотистих основ. За законами генетичного кодування різна інформація не може бути записана однаково, тому організми з неоднаковим нуклеотид- ним складом ДНК будуть різними. Якщо нуклеотидний склад двох організмів, які порівнюються, однаковий, то можлива як подібність, так і відмінність цих організмів, тому що генетичне кодування базується не лише на визначеному вмісті основ в одиниці кодування (триплеті), а й на їхньому взаємному розташуванні.
З таксономічною метою порівнюють молярний вміст суми гуаніну і цитозину (G+C) у відсотках від загальної кількості основ ДНК різних об'єктів. Виявлено, що вміст GC як у рослин, так і у тварин варіює у вузьких межах - 35-40 мол%, у той час як у бактерій - від 25 до 75 %. Унікальним виявилося те, що середній вміст GC-основ у штамів одного виду дуже близький або майже ідентичний. Така закономірність
доведена для багатьох груп бактерій, у тому числі і для представників роду Pseudomonas (табл. 7.2). У межах роду діапазон таких коливань теж досить обмежений і не перевищує 10-15 %. Така закономірність дозволяє використовувати значення вмісту GC-основ для упорядкування класифікації деяких таксономічних груп бактерій.
Проте не завжди подібний вміст G+C свідчить про спорідненість організмів. Наприклад, Pseudomonas aeruginosa і Mycobacterium phlei містять 60 % GC. Тому для більш точного визначення подібності (спорідненості) організмів використовують інші методи. Більші надії в систематиці прокаріотів покладають на метод гібридизації ДНК, який дозволяє кількісно оцінити ступінь гомологічності ДНК організмів, які порівнюються. Метод базується на здатності денатурованої (одноланцюгової) ДНК за певних умов денатурувавтися, тобто з'єднуватися з комплементарною ниткою й утворювати дволанцюгову молекулу. Кількість дволанцюгової ДНК, що утворилася внаслідок взаємодії одноланцюгових ДНК, виділених з різних організмів, є показником їхньої подібності.
Таблиця 7.2. Вміст G+C у різних штамів деяких видів роду Pseudomonas
Види |
Вміст G+C, мол% |
P. aeruginosa |
67,2 ±1,1 |
P. acidovorus |
66,8 ± 1,0 |
P. testosteroni |
61,8 ± 1,0 |
P. putida |
62,5 ± 0,9 |
Окрім аналізу молекул ДНК для встановлення ступеня спорідненості між прокаріотичними організмами розроблено методичні підходи, що дозволяють порівнювати продукти окремих генів, які виконують у клітині однакові функції. Це можуть бути білки (фередоксини, цитохроми тощо) або рРНК. Вибір рРНК для вирішення проблеми еволюційної систематики прокаріотів є вдалим через ряд причин: ці молекули виявлені в усіх клітинних форм життя; їхні функції завжди однакові; первинна структура в цілому характеризується високою консервативністю. Відомо, що в рибосомах прокаріотів та еукаріотів наявні три типи рРНК, які різняться молекулярною масою і коефіцієнтом седиментації. Інформаційна ємність великих молекул більша, але їх складніше аналізувати. Тому найбільш вдалою виявилась молекула 16 S рРНК (у прокаріотів) і 18 S рРНК (в еукаріотів). На базі отриманих результатів розраховано коефіцієнти подібності організмів, які порівнюються, і виявлено не дві групи організмів, що різняться прокаріотичним та еукаріотичним типом клітинної організації, а три (табл. 7.3). Одну групу утворюють всі еукаріоти: вищі рослини, тварини, дріжджі, водорості, гриби тощо. До цієї групи не увійшли органели еукаріотів (мітохондрії, хлоропласти). До другої групи, яка отримала назву справжні бактерії, належить переважна більшість прокаріотів. До неї ж потрапили на базі ступеня гомології 16 S рРНК мітохондрії і хлоропласти еукаріотів. До третьої групи увійшли прокаріоти (в основному мешканці екстремальних умов), які побудовані з макромолекул, більшість з яких є унікальними і не синтезується ні еукаріотами, ні бактеріями. Ця група отримала назву архебактерії. Згідно з даною ознакою запропоновано ділити всі живі організми на три над- царства або домени (домініони): Eukaryota, Eybacteria, Archaebactena.
Таблиця 7.3. Основні ознаки архебактерій, еубактерій та еукаріотів
Ознака |
Архебактерії |
Еубактерії |
Еукаріоти |
Типові організми |
Метаногени, екстремальні термофіли, галофіли |
Див. основні групи (за Bergey's Manual of Systematic Bacteriology, 1984) |
Протисти, гриби, рослини, тварини |
Типові розміри |
0,5-4,0 мкм |
0,5-4,0 мкм |
<5,0 мкм |
Геном |
Кільцева хромосома |
Кільцева хромосома |
Ядро з багатьма складними хромосомами |
Гістони |
Є |
Немає |
Є |
Зворотна гіраза |
Є |
Немає |
Немає |
Клітинна стінка |
Протеїн, псевдомуреїн |
Муреїн, ліпополісахарид |
Різна |
Мембрана |
Етерифіковані ізопреноїди |
Ефіри жирних кислот і гліцерину |
Ефіри жирних кислот, стероли |
Внутрішньоцитоплазматичні мембрани |
Немає |
Як правило нема або білкові |
Звичні для компартменталізації клітини |
Рибосоми |
70 S |
70 S |
80 S (в цитоплазмі) + 70 S рибосоми органел |
РНК-полімераза |
Складна |
Проста |
Складна |
Синтез білка інгібується: |
|||
Хлорамфеніколом |
Ні |
Так |
Ні |
Циклогексамідом |
Ні |
Ні |
Так |
1-го січня 1980 р. розроблено Міжнародний Кодекс номенклатури бактерій - МКНБ (International Code of Nomenclature of Bacteria - ICNB)
Згідно з МКНБ нові назви таксонів мають бути ефективно опубліковані в загальнодоступних виданнях, але новий таксон вважається дійсно визнаним, якщо його опис зроблено в International Jornal of Systematic and Evolution Microbiology (IJSEM) у вигляді окремої статті. Неопубліковані належним чином нові таксони мають подаватися в лапках.