МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
РОЗДІЛ 7. СИСТЕМАТИКА МІКРООРГАНІЗМІВ
КОРОТКА ХАРАКТЕРИСТИКА СИСТЕМАТИЧНИХ ГРУП ПРОКАРІОТІВ
Група 1. Спірохети. Група має порядок Spirochaetales із двома родинами: Spirochaetaceae і Leptospiraceae. Родина Spirochaetaceae має чотири роди: Spirochaeta, Cristispira, Treponema і Borrelia. Родина Leptospiraceae представлена лише одним родом - Leptospira. Це невелика група бактерій, які легко відрізняються від інших грамнегативних форм за морфологічною будовою клітини. Клітина спірохет сильно витягнута в довжину, спіральної форми, покрита еластичною клітинною стінкою, з одним або декількома витками спіралі. Спірохети характеризуються унікальним типом руху. Клітина має протоплазматичний циліндр, у якому локалізовані одна або декілька пар осьових ниток (фібрил). Кожна нитка відходить від диска, розташованого на полюсі клітини (циліндра). Скорочення фібрил супроводжується появою на поверхні клітини вторинної хвилі, що викликає її переміщення по субстрату.
Розмножуються спірохети поперечним поділом клітини. Серед них є аероби (Leptospira interrogans), факультативні анаероби (Spirochaeta aurantia) та анаероби (S. litoralis). За типом живлення - це хемоорганотрофи. Розміри клітини коливаються в широких межах. Великі клітини (до 250 мкм завдовжки) мають спірохети, які належать до родів Spirochaeta і Cristispira. Короткі клітини мають спірохети родів Treponema (5,020,0 мкм), Borrelia (3,0-10,0 мкм) і Leptospira (4,0-8,0 мкм).
Представники родів Treponema, Borrelia і Leptospira є паразитами людини та інших хребетних: Treponema pallidum - збудник сифілісу; T. pertenuae - збудник фрамбезії, захворювання, яке нагадує сифіліс (зустрічається в Центральній Америці, тропічній Африці та Індонезії); Borrelia recurrentis - збудник зворотної лихоманки (тифу) людини; Leptospira interrogans - збудник лептоспірозу, гострого інфекційного захворювання людини, сільськогосподарських, диких і промислових тварин, гризунів.
Група 2. Аеробні/мікроаерофільні, рухомі, спіралеподібні/віб- ріоїдні грамнегативні бактерії. Це досить гетерогенна група бактерій вібріоїдної або спіральної форми. У деяких видів при старінні культури частина клітин перетворюється в кокоподібні тіла з тонкою клітинною стінкою. Для представників роду Azospirillum характерне утворення великих цист. Клітини молодих культур забарвлюються за Грамом негативно. У старих культурах клітини представників роду Azospirillum можуть забарвлюватися позитивно. При вирощуванні в рідких поживних середовищах клітини мають джгутики, розташовані на одному полюсі (Azospirillum, Bdellovibrio, Cellvibrio, Vampirovibrio), обох полюсах (Aquaspirillum, Spirillum) або одному чи обох полюсах бактеріальної клітини (Campylobacter, Helicobacter). За типом живлення - хемоорганогетеротрофи, аероби або мікроаерофіли. Представники родів Azospirillum та Herbaspirillum у мікроаерофільних умовах здатні фіксувати молекулярний азот. Клітини Aquaspirillum magnetotacticumпроявляють таксис до магнітного поля.
Бактерії цієї групи виявляються в ґрунтах, прісних і солоних водоймах, на кореневій системі рослин, у кишковому тракті, ротовій порожнині людини та тварин. Представники родів Bdellovibno, Micavibrio і Vampirovibrio є хижаками щодо інших мікроорганізмів. Так, Bdellovibno bacteriovorus (лат. bdello - п'явка, vibrio - вібріон, тобто п'явко-вібріон) має один складний полярний джгутик (завтовшки до 50 мкм), який забезпечує досить швидкий рух клітини. За морфологічними і фізіологічними характеристиками бделовібріони мають двостадійний життєвий цикл, що складається зі стадії хижацтва, під час якої клітини не діляться, і репродуктивної стадії, що проходить у клітинах хазяїна. B. bacteriovorusприкріплюється до клітини хазяїна, втрачає джгутик, проникає крізь клітинну стінку (виділяючи літичні ферменти, які лізують клітинну стінку) у клітину-жертву і там розмножується. Цикл внутрішньоклітинного розвитку триває 3-5 год, потім клітини-паразити виходять із лізованої клітини-жертви і/або повторюють паразитичну фазу або утворюють цисти як форму спокою - дрібні, овальні чи сферичні клітини, покриті товстою оболонкою.
Бделовібріони лізують переважно грамнегативні бактерії, і насамперед псевдомонади та ентеробактерії.
Значний інтерес становлять представники роду Helicobacter - клітини у вигляді прямих чи зігнутих паличок або спіральної форми (0,5-1,0 х 2,55,0 мкм). Швидкий рух цих бактерій забезпечується численними джгутиками з чохлом, які розташовані на одному чи обох полюсах клітини, а інколи латерально. Мікроаерофіли. В атмосфері повітря, збагаченого 10 % СО2, і в анаеробних умовах ріст варіабельний. Усі штами швидко гідролізують сечовину. Ці бактерії було виділено зі слизової оболонки шлунка ссавців. Із наявністю деяких представників роду Helicbacter пов'язують захворювання гастритом та виразковою хворобою шлунка і 12-палої кишки, тому в останні роки змінилася стратегія лікування виразки. Типовим видом роду Helicobacter є H. pylori.
Група 3. Нерухомі (або рідко рухомі) грамнегативні зігнуті бактерії. Представники даної групи мають зігнуті, S-подібні, спіральні або кільцеподібні клітини. У деяких випадках (бактерії роду Spirosoma) кінці клітин можуть перекриватися, утворюючи кільце із зовнішнім діаметром 1,5-3,0 мкм, а довжина синусоїдально зігнутої нитки досягає 50 мкм. Інколи дугоподібно зігнуті клітини розташовуються у вигляді листка конюшини (Branchyarcus), деякі утворюють скупчення рухомих клітин із джгутиками (Pelosigma). Можуть проявляти дихальний (Runella) або бродильний (Meniscus) тип метаболізму. Хемоорганотрофи. Облігатні аероби (Branchyarcus thiophilus), аеротолерантні анаероби (Meniscus glaucopis). Виявляються в ґрунтах, прісних та солоних водоймах.
Група 4. Грамнегативні аеробні/мікроаерофільні палички та коки. Це досить гетерогенна група мікроорганізмів, але всі вони мають дихальний тип метаболізму і використовують як кінцевий акцептор електронів кисень. Майже всі представники (за винятком мікроа- ерофілів) ростуть в атмосфері повітря (21 % О2). Деякі здатні до анаеробного дихання з використанням нітрату, фумарату або інших кінцевих акцепторів електронів. Бактерії кількох родів можуть фіксувати молекулярний азот.
До родини Pseudomonadaceae належать представники родів Pseudomonas, Xanthomonas, Xanthobacter. Хемоорганотрофи, деякі види факультативні хемолітотрофи. Бактерії цієї родини значно поширені в ґрунті, прісних та солоних водоймах, у мулі та стічних водах, де викликають інтенсивні процеси мінералізації органічних речовин. Використовуються як продуценти біологічно активних речовин: органічних кислот (глюконової, α-кетаглутарової, піровиноградної), амінокислот (аспарагінової, валіну, глутаміну), ферментів (аспарагінази, пероксидази). Серед представників цієї родини є фітопатогени (Xanthomonas) і патогени для людини та тварин (Pseudomonas aeruginosa - синьогнійна паличка, яка може спричинити менінгіт, отит, пневмонію, післяопераційні нагноєння). Деякі види мають ентомопатогенні властивості та виявляють антибіотичну активність щодо бактерій, дріжджів та грибів.
До родини Azotobacteriaceae належать види, клітини яких мають тенденцію до поліморфізму - зміни морфології клітини залежно від фізіологічного стану культури та умов культивування. Деякі види утворюють цисти. Це облігатні аероби, хемоорганогетеротрофи, які виявляються в ґрунтах, на поверхні рослин та у водоймах. Характерною особливістю родини є здатність її представників фіксувати молекулярний азот. Azotobacter chroococcum - перший вільноживучий аеробний фіксатор молекулярного азоту, описаний М. Бейєринком у 1901 р. Клітини овальної форми, плеоморфні - від паличкоподібних до коків. Розташовуються парами або утворюють короткі ланцюжки, оточені загальною капсулою. При старінні утворюються цисти (форма спокою). Представники роду Azomonas (A. agilis) цист не утворюють.
Родина Rhizobiaceae у 9-му виданні "Bergey's Manual of Systematic Bacteriology" представлена двома родами: Rhizobium та Agrobacterium, але останнім часом описано й офіційно визнано нові роди бульбочкових бактерій (Azorhizobium, Bradyrhizobium тощо). Рід Rhizobium об'єднує симбіотичні фіксатори N2. Це грамнегативні, хемоорганотрофні, рухомі (мають один полярний чи субполярний джгутик або перитри- хальне джгутикування) паличкоподібні бактерії, особливістю яких є здатність викликати розростання тканин кореневої системи з утворенням бульбочок і фіксувати азот атмосфери, перебуваючи в симбіотичних зв'язках із бобовою рослиною (рис. 7.6). В основу класифікації бульбочкових бактерій покладено їхню специфічність щодо рослини- хазяїна: R. leguminosarum b. viceae (бактерії гороху, вики, кормових бобів), R. leguminosarum b. trifolii (конюшини), R. leguminosarum b. phaseoli (квасолі), R. loti (люпину), R. meliloti (люцерни), R. galegae (козлятника). У тропічних і субтропічних широтах виявляються представники роду Bradyrhizobium (B. japonicum). Бактерії роду Agrobacterium - це пухлиноутворювальні фітопатогени (за винятком A. radiobacter). Вони викликають розростання тканин рослин у вигляді галів на кореневій системі та стеблах рослин - гал корончастий (бактеріальний), корінь волосяний, рак стебла бактеріальний. Індукція пухлин корелює з наявністю у клітинах бактерій великої пухлиноіндукуючої плазміди (Ti-плазміда). Типовий вид - A. tumefaciens.
Родина Methylococcaceae об'єднує бактерії, які використовують як єдине джерело вуглецю одновуглецеві органічні сполуки - метан або метанол. Представники родів Methylomonas (M. methanica, рухомі) і Methylococcus (M. capsulatus, нерухомі) відрізняються за морфологією та здатністю до руху.
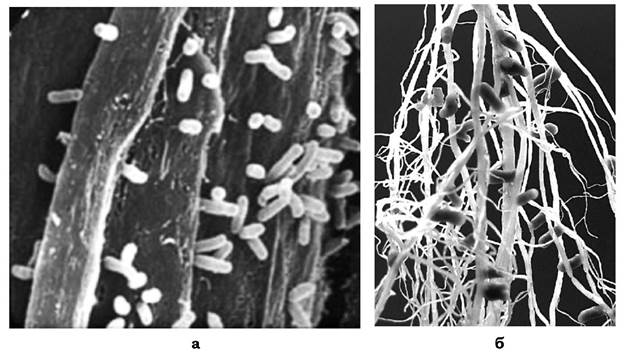
Рис. 7.6. Бульбочкові бактерії роду Rhizobium:
а - клітини Rhizobium, що проникають до рослинної тканини; б - сформовані бульбочки на бобовій рослині
Родина Halobacteriaceae об'єднує галотолерантів роду Halomonas (H. elongata), які здатні розвиватися за наявності в середовищі 0,0520 % NaCl. Метаболізм дихального типу, але деякі представники здатні до росту в анаеробних умовах з використанням нітратів як кінцевого акцептора електронів. Виділені з обладнання для добування грубої солі.
Характерною особливістю представників родини Acetobacteriaceae є здатність окиснювати спиртові розчини з утворенням оцтової кислоти. Клітини паличко- чи еліпсоподібної форми або зігнуті, розташовуються поодиноко, парами або утворюють ланцюжки клітин. Це облігатні аероби, рухомі (джгутикування перитрихальне чи латеральне) або нерухомі організми. Хемоорганотрофи, які виявляються у вині, сидрі, кефірі та ін. Родина представлена двома родами: Acetobacter (A. aceti) і Gluconobacter (G. oxydans).
До родини Neisseriaceae віднесено чотири роди: Neisseria, Moraxella, Kingella, Acinetobacter. Представники перших трьох родів - паразити слизових оболонок: Neisseriagonorrhoeae - збудник гонореї, N. meningitidis - збудник менінгіту, Moraxella lacunata та Kingella kingae - паразити слизових оболонок людини і теплокровних тварин. Ці бактерії потребують для свого розвитку складних поживних середовищ. Рід Acinetobacter представлений сапрофітними формами, які значно поширені у природі.
Група 5. Факультативно анаеробні грамнегативні палички. Ця група об'єднує представників трьох родин: Enterobacteriaceae, Vibrionaceae і Pasteurellaceae. До родини Enterobacteriaceae віднесено рухомі (перитрихи), аспорогенні, хемоорганотрофні, сапрофітні та патогенні організми. Метаболізм дихального або бродильного типу. Поширені в ґрунті, воді, виявляються на овочах, рослинах, в організмі людини і тварин. Певні серотипи E. coli, представники родів Klebsiella, Enterobacter, Proteus, Providencia можуть спричиняти внутрішньолікарняні інфекції. Типовий рід - Escherichia, типовий вид - E. coli (кишкова паличка). Представники цього роду добре ростуть на звичайних поживних середовищах при температурі 37 0С з утворенням колоній двох типів: S-колонії (гладенькі) - слабкоопуклі, вологі з блискучою поверхнею і рівним краєм, сіруваті, добре емульгуються в сольовому розчині; R-колонії (шорсткі) - сухі, не емульгуються в сольовому розчині. Для диференціації використовують середовище Ендо, на якому кишкова паличка утворює рожеві колонії з металевим блиском, та серологічні реакції. До роду Escherichia належить також E. blattae.
Близькими до E. coli є коліформні бактерії родів Salmonella, Shigella і Citrobacter. До роду Salmonella належать види, які можуть бути збудниками черевного тифу (S. typhi), паратифу (S. paratyphi) та внутрішньолікарняних сальмонельозів (S. typhimurium). Шигели - збудники бактеріальної дизентерії: S. dysenteriae, S. flexneri, S. sonnei.
Окремі види роду Proteus можуть спричинити післяопераційні ускладнення. Фітопатогенні види зустрічаються серед бактерій родів Erwinia, Serratia, Hafnia та Enterobacter. Бактерії роду Yersinia мають овальні або паличкоподібні клітини, рухомі, але їхня рухомість залежить від температури культивування. Y. pestis (збудник чуми) був описаний у 1894 р. Йєрсеном.
До родини Vibrionaceae належать прямі або зігнуті рухомі палички, які виявляються у водоймах, на поверхні та в організмі водних тварин. Бактерії роду Aeromonas - мешканці прісних водойм, вони можуть викликати захворювання жаб, риб або інфікувати організм людини, що супроводжується явищами діареї та бактеріємії. Vibho cholerae - збудник холери - гострого інфекційного захворювання з великою тенденцією до епідемічного поширення. Більшість штамів бактерій роду Photobacterium ростуть у мінеральному середовищі, виготовленому на основі морської води за наявності D-глюкози і NH4CI. Два види цього роду (P. leiognathi, P. phosphoreum) здатні до біолюмінесценції. Ці бактерії
можна виділити з морських риб. Якщо морську рибу в неглибокому посуді залити солоною водою (~ 3 % NaCl) і залишити на декілька діб у холодильнику, то на поверхні риб з'являються колонії, які світяться.
Родина Pasteurellaceae представлена плеоморфними, нерухомими, аеробними або факультативно анаеробними бактеріями. Клітини - від кокоподібних до прямих паличок, які можуть роздуватися або формувати ниткоподібні структури. Хемоорганотрофи з дихальним і бродильним типами метаболізму. Це паразити хребетних, переважно ссавців і птахів. Pasteurella multocida - збудник геморагічної септицемії великої рогатої худоби, холери птахів і пневмонії сільськогосподарських тварин. Haemophilus influenzae - основний збудник менінгіту в дітей, також може бути причиною запалення середнього вуха, хронічного бронхіту, пневмонії. Цей вид уперше було описано в 1889 р. як етіологічний фактор грипу. H. ducreyi - збудник м'якого шанкру, або шанкроїду.
Група 6. Грамнегативні, анаеробні, прямі, зігнуті й спіральні бактерії. Це облігатні анаероби, хемоорганотрофи, нерухомі або рухомі палички, які тяжіють до плеоморфізму. Бактерії різних родів відрізняються за кінцевими продуктами бродіння: Butyrovibrio - з глюкози утворюють бутират; Succinovibdo - сукцинат, ацетат, форміат та ін.; Fusobactedum - масляну кислоту; Leptotrhichia - молочну кислоту; Bacteroides - суміш кислот (янтарної, оцтової, мурашиної та ін.). Бактерії цієї групи виявляються в травному тракті людини, тварин (Selenomonas sputigena - мешканець ротової порожнини людини; S. ruminantium - рубця жуйних). Окремі види Fusobactedum виділяються з крові та осередків нагноєння.
Група 7. Бактерії, які здійснюють дисиміляційне відновлення сульфату або сірки. До цієї групи належать хемолітотрофні облігатні анаероби. Сульфатредукуючі бактерії відновлюють сульфат або сірку до H2S. Донорами електронів служать H2 або органічні сполуки. Окиснення органічних сполук може бути неповним (з утворенням ацетату як кінцевого продукту) або повним (з утворенням СО2). У багатьох видів донорами електронів виступають Н2, лактат, жирні кислоти, етанол або дикарбонові кислоти. Можливий автотрофний ріст за рахунок асиміляції Н2, СО2 і сульфатів. Типові місця виявлення - безкисневі осади або придонний шар у прісних, морських чи засолених водоймах. Термофільні види зустрічаються в гарячих джерелах та підводних вивідних каналах гідротерм. Група гетерогенна за своїми морфологічними, фізіологічними та біохімічними ознаками. Термофільні сульфатредукуючі архебактерії та екстремально термофільні архебактерії віднесено до груп 32 і 35 відповідно. До цієї групи належать представники декількох родів: Desulfolobus, Desulfomicrobium, Desulfomonas, Desulfovibrio, Thermodesulfobacterium.
Група 8. Анаеробні грамнегативні коки. Коки діаметром від 0,3-0,5 до 2,5 мкм. Розташовуються парами, поодиноко, формують нехарактерні угруповання або ланцюжки нерухомих клітин. Грамнегативні, але мають тенденцію стійкості до знебарвлення. Хемоорганотрофи, вибагливі до поживних речовин. Представники родів Acidaminococcus, Magasphaera та Veillonella мають метаболізм бродильного типу. Для деяких родів характерне зброджування лактату з утворенням СО2, Н2 і різних летких жирних кислот з 2-6 атомами вуглецю. Види роду Syntrophococcus мають дихальний тип метаболізму і потребують для свого росту наявності в середовищі цукрів як донорів електронів, а також форміату як екзогенної електрон-акцепторної системи. Виявляються переважно в організмі жуйних тварин, гризунів, свиней, а також людини. Magasphaera cerevisiae зустрічається в пиві при тривалому його зберіганні.
Група 9. Рикетсії та хламідії. Поділяються на два порядки: Rickettsiales і Chlamidiales. Рикетсії представлені організмами, які мають клітини паличкоподібної форми, грамнегативні, нерухомі, розмножуються бінарним поділом усередині клітин хазяїна, де паразитують, не ростуть на звичайних поживних середовищах. Але кожна з цих ознак може мати певні винятки: деякі рикетсії рухомі (мають один джгутик); у фарбованих препаратах мають кільцеподібну форму; можуть забарвлюватися за Грамом позитивно. Паразитичні види рикетсій асоціюються з ретикулоендотеліальними клітинами й клітинами ендотелію судин або еритроцитів крові хребетних. За несприятливих умов здатні утворювати форми спокою і дегенеративні форми. Форма спокою являє собою клітину з ущільненою клітинною стінкою і цитоплазмою. На підставі специфічності існування в організмі хазяїна їх поділяють на три родини: Rickettsiaceae, Anaplasmataceae, Bartonellaceae.
До родини Rickettsiaceae належать внутрішньоклітинні паразити, які поділяються на три триби: Rickettsieae (роди Rickettsia, Rochalimaea, Coxiella) - патогенні форми; Erlichieae(роди Erlichia, Cowdria, Neorickettsia) - непатогенні для людини види; Wolbachieae (роди Wolbachia, Rickettsiella) - представлена непатогенними для хребетних видами, які не культивуються на безклітинних середовищах.
Рикетсії триби Rickettsieae є збудниками багатьох небезпечних для людини захворювань: висипного тифу (R. typhi, R. prowazekii), кліщових плямистих лихоманок (R. sibirica), лихоманки Q (англ. query - неясний), збудником якої є Coxiella burnetii.
Родина Anaplasmataceae представлена облігатними паразитами, які виявляються в еритроцитах, на поверхні еритроцитів або у плазмі крові диких і свійських хребетних - корів, кіз, оленів (роди Anaplasma, Aegyptianella, Haemobartonella, Eperytozoon).
Рикетсії родини Bartonellaceae мають паличкоподібні, сферичні або кільцеподібні клітини, які виявляються в еритроцитах або на поверхні еритроцитів людини, у клітинах тканин (рід Bartonella), а також в еритроцитах тварин (рід Grahamella).
Порядок Chlamidiales має родину Chlamidiaceae і рід Chlamidia. Це кокоподібні мікроорганізми (0,2-1,5 мкм) з характерним внутрішньоклітинним способом розмноження. Першим етапом інфекційного процесу є адсорбція елементарного тільця (ЕТ) хламідій на плазмалемі чутливої клітини організму хазяїна. Елементарні тільця - це інфекційна форма хламідій розміром 0,2-0,4 мкм, які не здатні до розмноження, але здатні до інфікування (проникнення) клітини-хазяїна. Проникнення хламідій відбувається шляхом ендоцитозу протягом 710 год. При цьому ЕТ при проникненні в клітину використовує для росту клітинну АТФ і через 6-8 год перетворюється на ретикулярне тільце (РТ). Ретикулярні тільця (неінфекційна форма) - це великі (0,81,5 мкм) комірчасті сфероїди, здатні до активного ділення. РТ утворюють мікроколонії - хламідійні включення (тільця Гальберштадтера - Провачека). Протягом 18-24 год вони локалізовані в цитоплазматичному пухирці, утвореному з мембрани клітини хазяїна. У мікроколоніях може міститися від 100 до 500 клітин. Процес визрівання (проміжні тільця) і трансформація РТ шляхом ділення в ЕТ триває 3642 год. Цикл завершується руйнуванням ураженої клітини. Хламідії можуть вивільнятися з інфікованої клітини, зберігаючи життєздатність клітини-хазяїна (при виході ЕТ шляхом екзоцитозу), що призводить до безсимптомного перебігу захворювання (рис. 7.7).
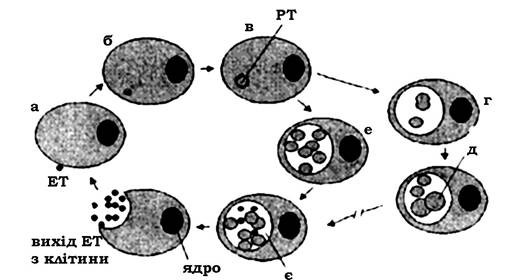
Рис. 7.7. Життєвий цикл хламідій:
а - адсорбція елементарного тільця (ЕТ) хламідії на плазмалемі еукаріотичної клітини; б - проникнення ЕТ всередину клітини; в - трансформація ЕТ та РТ; г - обмежене ділення; д - абератні форми хламідій; е - розмноження шляхом бінарного ділення; є - трансформація РТ в ЕТ
Під час вивчення механізмів персистенції хламідій було отримано дані, згідно з якими в цитоплазматичних включеннях, окрім повноцінних інфекційних (елементарних) тілець, виявляються анормальні форми: гігантські форми, у тому числі сферопластоподібні тіла, діаметром до 5000 нм; дрібні, позбавлені клітинної стінки структури розміром 100-250 нм; пухирцеподібні форми діаметром 30-80 нм.
Вважається, що за деякими ознаками хламідії займають проміжне положення між бактеріями та вірусами. Комплекс бактеріоподібних властивостей хламідій забезпечує: збереження морфологічної суті протягом усього життєвого циклу; поділ вегетативних форм; наявність клітинної стінки грамнегативного типу бактерій; наявність ДНК, РНК, ферментативної активності; здатність забарвлюватися аніліновими барвниками; чутливість до антибіотиків широкого спектра дії; наявність родоспецифічного антигену, загального для всіх хламідій.
З вірусами їх зближує: дрібні розміри (вони фільтруються через бактеріальні фільтри); нездатність до росту на штучних поживних середовищах (безклітинних субстратах); здатність до тривалої персистенції в організмі хазяїна, прояв енергетичного паразитизму.
Хламідії - паразити людини, ссавців, птахів; вони викликають ряд захворювань у людини: трахому, венеричну лімфогранулому, інклюзій - ний кон'юнктивіт (C. trachomatis), орнітоз (C. psittaci). У 1988 р. Юридична комісія Міжнародної асоціації мікробіологічних товариств офіційно визнала третій вид хламідій - C. pneumoniae, який викликає респіраторну патологію людини. У 1990 р. японські мікробіологи описали четвертий вид - C. pecorum, з яким пов'язують розвиток патології суглобів. А в 2000 р. було затверджено нову міжнародну класифікацію хламідій, яка базується на наявності > 95 % гомології в нуклеотидній послідовності генів 16S і 23S рРНК для всіх представників роду і > 90 % - родини. Згідно з новою класифікацією до порядку Chlamydiales увійшло чотири родини: Chlamydiaceae, Parachlamydiaceae, Simkaniaceae, Waddliaceae. Найрадикальніші зміни відбулися в систематиці родини Chlamydiaceae, в якій на даний момент виділено два роди: Clamydia (C. trachomatis, C. suis, C. mundarum) i Clamydophila (C. pneumoniae, C. pecorum, C. psittaci, C. abortus, Ccaviae, C. felis). Інші три родини включають по одному роду з єдиним представником: Parachlamydia (P acanthamoebae); Simkania (S. nagavensis); Waddlia (W. chondrophila).
Група 10. Аноксигенні фототрофні бактерії. Фотосинтезуючі бактерії, які характеризуються специфічним набором пігментів (бактеріохлорофіли і каротиноїди) та особливим типом фотосинтезу, що не супроводжується виділенням кисню (аноксигенний). Морфологічно це гетерогенна група - коки, палички, звивисті форми, нерухомі й рухомі. Клітини розташовуються поодиноко або в угрупованнях правильної чи неправильної форми. Зустрічаються також одноклітинні або однорядні багатоклітинні нитчасті форми. Довжина клітини коливається в межах 1,0-5,0 мкм (інколи досягає 100 мкм), а її товщина - 0,3-6,0 мкм. Ендоспор не утворюють, грамнегативні, розмножуються бінарним поділом, деякі види (Rhodopseudomonas palustris, R. viridis) - брунькуванням.
Фотосинтетичні пігменти можуть бути локалізовані в: ЦПМ (Heliobacillus - мають джгутики, Heliobacterium - мають ковзний тип руху); цитоплазматичних мембранних структурах (Amoebobacter, Chromatium, Thiospirillum); хлоросомах (Chlorobium, Ancalochloris, Chloroflexus, Chloronema).
Енергетичний метаболізм більшості видів проходить при освітленні (фотосинтез), але в деяких видів може відбуватися в темряві або за наявності кисню (окиснення, Thiocapsa), чи в анаеробних умовах під час ферментативного перетворення органічних сполук (деякі пурпурові бактерії).
Група 11. Оксигенні фототрофні бактерії. Ця група представлена ціанобактеріями і прохлорофітами.
Ціанобактерії (раніше відомі як синьозелені водорості) - сферичні, паличкоподібні, зігнуті одноклітинні, колоніальні і багатоклітинні (нитчасті), грамнегативні організми, які мають ригідну клітинну стінку, покриту капсулою, і ковзний тип руху (Chroococcales, Pleurocapsales, Oscillatoriales, Nostocales, Stigonematales). Розмножуються бінарним поділом, брунькуванням, гормогоніями, спорами, акінетами, уламками трихом. У тилакоїдах виявляється комплекс фотосинтетичних пігментів - хлорофіл а, алофікоціанін, фікоціанін, фікоерітрини.
Порядок FTochlorales (прохлорофіти) об'єднує одноклітинні або нитчасті (розгалужені чи нерозгалужені) прокаріоти, які нагадують ціанобактерії, але на відміну від останніх містять хлорофіли а та в і позбавлені додаткових червоних чи синіх білінових пігментів. Виявляються виключно як позаклітинні симбіонти в асоціаціях з колоніальними асцидіями в тропічних чи субтропічних морських середовищах.
Група 12. Аеробні хемолітотрофні бактерії і родинні організми. Ця група прокаріотів поділяється на три підгрупи: 1) безбарвні сіркоокиснювальні бактерії; 2) залізо- і манганоокиснювальні і/або бактерії, які їх осаджують; 3) нітрифікуючі бактерії.
Безбарвні сіркоокиснювальні бактерії представлені всіма відомими морфологічними формами. Здатні до росту при pH від 1,0 до 10,5. Гра- мнегативні, хемолітотрофи, які виявляються в місцях, де є відновлені сполуки сірки: ґрунт, на межі аеробних та анаеробних зон водойм, вивідних каналах гідротерм (Macromonas, Thiobacterium, Thiobacillus).
Залізо- і манганоокиснювальні і/або бактерії, що їх осаджують, - це гетерогенна група мікроорганізмів, які часто утворюють специфічні агрегати. У таких агрегатах визначаються структури, забарвлені оксидами заліза або мангану. Клітина паличкоподібна (Thiobacillus), паличка зігнута (Leptospirillum), коки неправильної форми (Sulfolobus). Клітина може мати вирости у вигляді простек (простекобактерії - Hyphomicrobium) або стеблинок (стеблинкові бактерії - Gallionella). Зустрічаються нитчасті бактерії з чохлами, просоченими оксидами (Lepthothrix). За Грамом забарвлюються позитивно чи негативно або не мають клітинної стінки. Це мікроаерофіли, але деякі з них ростуть при звичайних парціальних тисках О2.
Залежно від субстрату, який окиснюється, нітрифікуючі бактерії поділяються на дві секції: секція А - бактерії, які окиснюють нітрит (Nitrobacter, Nitrospina, Nitrococcus, Nitrospira) і секція В - бактерії, які окиснюють аміак (Nitrosomonas, Nitrosococcus, Nitrosospira,
Nitrosolobus, Nitrosovibrio).
Група 13. Бактерії, які брунькуються і/або мають вирости.
До цієї групи належать бактерії незвичної форми з виростами та складними клітинними циклами (рис. 7.8).
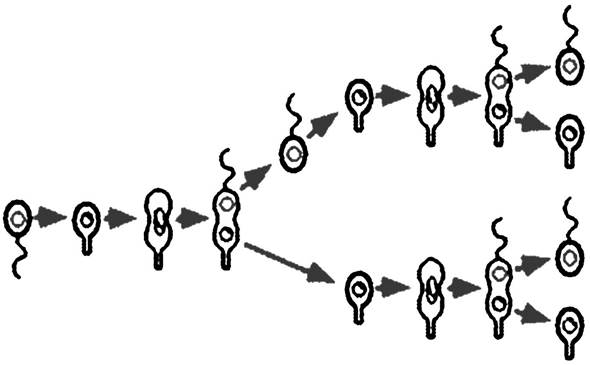
Рис. 7.8. Стадії життєвого циклу Hyphomicrobium
У межах групи виділяють три підгрупи.
Підгрупа 1. Простекобактерії - це одноклітинні бактерії, які утворюють одну або більше простек на клітину. Розмноження відбувається шляхом брунькування або бінарного поперечного поділу. Клітини без простек - паличкоподібні, вібріоїдні, кокоподібні або тетраедричні. Простеки можуть надавати клітині форму булави, зірки, спіралі. Гетеротрофи, більшість представників - аероби (Caulobacter, Dchotomicrobium, Hyphomicrobium, Pedomicrobium, Prosthecomicrobium, Stella), деякі - факультативні анаероби (Ancalomicrobium). Олігокарбофіли, добре ростуть на розведених середовищах. Одні види краще ростуть за наявності дріжджового екстракту, інші - потребують вітаміни. Виявляються у водоймах та ґрунтах.
Підгрупа 2. Порядок Planctomycetales - це одноклітинні бактерії, які утворюють розетки або нитки. Клітини кокоподібні, овальні або грушоподібні. Розмножуються брунькуванням, можуть утворювати вирости, які називаються стеблинками. Виявляються в прісних, морських та інших солоних водоймах (Planctomyces).
Підгрупа 3. Бактерії, які брунькуються і/або мають вирости. Так, представники роду Nevskia мають довгі (1,0-6,0 х 3,0-12,0 мкм), паличкоподібні клітини з прозорими стеблинками. Стеблинки - це капсульна речовина, яка виділяється з одного боку клітини. Після завершення бінарного поперечного поділу дочірні клітини утримуються разом стеблинками, утворюючи Y-подібні гілки. У природних середовищах вони виявляються у вигляді агрегатів, які нагадують кольорову капусту (N. ramosa).
Група 14. Бактерії, які мають чохол. Ростуть у вигляді ланцюжків клітин - ниток, завширшки 0,4-7,0 мкм. Грамнегативні. Нитки ростуть у трубочках з позаклітинного матеріалу, який класифікується як чохол. Інколи чохол настільки тоненький, що його важко виявити при фазово-контрастній мікроскопії. Чохли в результаті осадження оксидів заліза й мангану можуть бути забарвленими. Бактерії, які виявляють здатність до руху, мають джгутики. Аероби. Хемоорганогетеротрофи. Деякі осаджують оксиди заліза та мангану в чохлі або на його поверхні, можуть окиснювати Fe(II) і Mn(II). Виявляються у водних середовищах. Представники роду Crenothrix мають клітини від циліндричних до дископодібних, діляться шляхом утворення поперечних перетинок і формують нитки завдовжки до 1 см у чохлах, які можуть прикріплюватися до щільного субстрату. Чохол може бути безбарвним на вершині або покритий оксидом заліза чи мангану в основі. У нитках можна спостерігати збільшення поперечного септу- вання на одному чи обох кінцях з утворенням сферичних клітин розмноження - "макрогонідій".
Група 15. Нефотосинтезуючі ковзні бактерії, що не утворюють плодових тіл. Група морфологічно гетерогенна - об'єднує бактерії у формі коків, спіралей, довгих гнучких паличок, покритих чи непокритих чохлом ниток, гнучких трихом і розеток, які здійснюють ковзний рух. Деякі представники можуть мати газові вакуолі, інші містять включення сірки. Бактерії роду Beggiatoa ростуть хемоавтотрофно, всі інші - хемогетеротрофно, деякі здатні до міксотрофного росту. Серед них є одноклітинні паличкоподібні (Cytophaga, Flexibacter, Sporocytophaga), плоскі нитчасті (Alysiella) та сіркоокиснювальні ковзні бактерії (Achromatium, Beggiatoa, Thiothrix).
Група 16. Ковзні бактерії, які утворюють плодові тіла: міксобактерії. Паличкоподібні грамнегативні бактерії, які здатні до ковзного руху, утворюють плодові тіла і стійкі до висушування міксоспори. Клітини (0,6-1,2 х 2,0-10,0 мкм) двох морфологічних типів: а) тонкі гнучкі палички зі звуженими кінцями; б) відносно товсті палички циліндричної форми із заокругленими кінцями. У результаті ковзного руху клітин колонії міксобактерій поширюються по поверхні субстрату, і тому вони мають назву швармів. Усередині шварма клітини розподілені нерівномірно, концентруючись у радіальних тяжах або в масивних складках по периферії шварма.
В умовах голодування клітини скупчуються або агрегують у певних ділянках шварма, утворюючи великі глобулярні або гребенеподібні маси, які містять 104-106 клітин. Ці маси диференціюються в плодові тіла, форма й будова яких залежить від виду (рис. 7.9). Плодові тіла можуть бути простими, без спорангіолей (Myxococcus, Angiococcus, Archangium), і складними, тобто складатися з кількох одиниць (Cystobacter, Stigmatella, Polyangium, Chondromyces). Розмір плодових тіл варіює від 100 до 600 мкм. Під час дозрівання плодового тіла вегетативні клітини перетворюються в міксоспори - форма спокою (стара назва - мікроцисти).
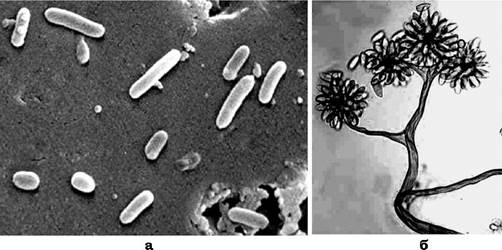
Рис. 7.9. Вегетативні клітини (а) і плодове тіло (б) Chondromyces crocatus
За типом живлення міксобактерії - хемоорганотрофи, здатні до деструкції різних біомакромолекул. Виявляються в ґрунтах, деревині, корі живих і відмерлих дерев, воді.
Група 17. Грампозитивні коки. Це численна і гетерогенна група мікроорганізмів, об'єднаних в одну на підставі таких загальних ознак, як сферична форма клітини і позитивна реакція при забарвлюванні за Грамом. Серед них є аероби (Micrococcus, Planococcus, Deinococcus), факультативні анаероби (Staphylococcus, Stomatococcus, Streptococcus, Leuconostoc, Pediococcus, Aerococcus), анаеробні роди (Peptococcus, Peptostreptococcus, Ruminococcus, Coprococcus, Sarcina).
Група має дві родини (Micrococcaceae і Deinococcaceae) і ряд родів, які займають незалежне систематичне положення.
До родини Micrococcaceae належать об'єкти з досить різноманітними харчовими потребами. Вони ростуть за умов вмісту в середовищі 5 % NaCl, а деякі штами здатні розвиватися за наявності 10 % NaCl. Деякі види патогенні для людини і тварин. До цієї родини віднесено роди Micrococcus (M. lylae, M. roseus, M. agilis, M. knstinae),Stomatococcus (S. mucilaginosus), Planococcus (P. citreus), Staphylococcus (S. aureus, S. epidermidis, S. haemolyticus, S. saprophyticus, S. warned).
До роду Staphylococcus належать бактерії, які мають сферичну форму клітини діаметром 0,5-1,5 мкм. У препаратах виявляються поодинокі клітини, інколи вони розташовуються парами або утворюють характерні угруповання (грона винограду), що є результатом поділу
клітини більш як в одній площині. У клітинних стінках визначаються як пептидоглікан, так і тейхоєві кислоти. Діамінокислотою пептидоглікану є L-лізин. Оптимальна температура росту - 35-40 0С, температурний діапазон росту - 15-45 0С. Оптимум pH - 7,0-7,5, межі pH - від 4,2 до 9,3. Представники деяких видів (S. aureus, S. cohnii, S. intermedius, S. epidermidis) проявляють гемолітичну активність.
До родини Deinococcaceae входить один рід - Deinococcus (D. radiodurans), клітини представників якого мають сферичну форму (діаметр 0,5-3,5 мкм), розташовуються парами, а частіше формують тетради. Нерухомі, хемоорганотрофи з дихальним типом метаболізму.
За такими ознаками, як здатність до росту при температурі 10 0С і 45 0С у середовищі з 6,5 % NaCl, при pH 9,6 у середовищі, що містить 40 % жовчі, спричиняти α- або β-гемоліз, гідролізувати аргінін, крохмаль і гіпурат, здатність до росту в анаеробних умовах, представників роду Streptococcus поділяють на декілька груп:
- піогенні стрептококи (S. pyogenes, S. agalactiae, S. equi, S. pneumoniae);
- стрептококи ротової порожнини (S. salivarius, S. sanguis, S. mutans, S. sobrinus);
- ентерококи (S. faecalis, S. faecium);
- молочнокислі стрептококи (S. lactis, S. raffinolactis);
- анаеробні стрептококи (S. hansenii, S. pleomorphus);
- інші стрептококи (S. thermophillus, S. bovis).
Варто зазначити, що представники таких фізіологічних груп, як ентерококи та молочнокислі стрептококи, отримали відповідні родові назви Enterococcus та Lactococcus і розглядаються паралельно у відповідних таксономічних групах. Тобто слід пам'ятати, що такі представники, як Streptococcus lactis та Enterococcus lactis, є ідентичними.
Представники роду Leuconostoc (L. mesenteroides, L. paramesenteroides, L. lactis) мають клітини сочевицеподібної форми, розташовуються парами або формують ланцюжки, нерухомі, хемоорганотрофи, які потребують багатих поживних середовищ. Оптимальна температура росту - 20-30 0С, але здатні рости в температурних межах 5-30 0С. На середовищах із сахарозою (α-глюкозил-β-фруктоза) продукують значну кількість слизу, чим завдають значних збитків під час виробництва цукру ("хвороба жаб'ячої ікри»).
До роду Sarcina належать представники, які мають майже сферичні клітини (1,8-3,0 мкм) і утворюють характерні угруповання - пакети, що містять 8, 16, 32... клітини. Нерухомі, пігменти не синтезують. Метаболізм бродильного типу, облігатні анаероби. S. ventriculi
виділяється з ґрунту, організму людей із хворим шлунком, поверхні насіння злакових. Клітина має додатковий зовнішній целюлозний шар. S. maxima виділяється з насіння злакових, клітина не має зовнішнього додаткового шару.
Група 18. Грампозитивні палички й коки, які утворюють ендоспори. Таксономічні зв'язки в межах цієї групи вивчені недостатньо. Клітини більшості представників мають паличкоподібну форму. Виняток становлять штами, віднесені до роду Sporosarcina; вони представлені кокоподібними клітинами, які здатні до утворення тетрад або пакетів. Товщина клітини більшості представників коливається в межах 0,3-2,0 мкм. Винятком є штами роду Oscillospira, діаметр клітин яких досягає 3,0-6,0 мкм клітини мають паличкоподібну форму або утворюють нитки, розділені перетинками на дископодібні клітини.
Більшість забарвлюється за Грамом позитивно, але можуть бути грамваріабельними або грамнегативними. Як правило, рухомі, з перитрихальним типом джгутикування. Метаболізм та екологія різних представників сильно варіює. До даної групи віднесено ряд родів, які займають незалежне систематичне положення: Amphibacillus, Bacillus, Clostridium, Desulfotomaculum, Oscillospira, Sporohalactobacter,
Sporolactobacillus, Sporosarcina, Sulfobacillus. За відношенням до кисню ендоспороутворювальні бактерії представлені всіма типами: облігатні аероби (Sporosarcina), аероби та факультативні анаероби (Bacillus), факультативні анаероби (Sporolactobacillus), облігатні анаероби (Clostridium, Desulfotomaculum, Oscillospira). Серед представників цієї групи є збудники маслянокислого (Clostridium butyricum) та аце- тонобутилового бродіння (C. acetobutylicum), уролітичні бактерії (Sporosarcina ureae, Bacillus pasteurii), вільноживучі фіксатори молекулярного азоту (C. pasteurianum), продуценти антибіотиків, зокрема, граміцидину С (B. brevis var. G.-B.), поліміксину (B. polymyxa), субтиліну (B. subtilis), а також ферментів - протеаз (B. subtilis), амілаз (B. licheniformis), амінокислот - L-глутамату (B. megaterium), L-триптофану (B. subtilis), ентомоцидних токсинів (B. thuringiensis) та інших біологічно активних речовин.
Щодо екології - це космополіти, які виявляються в ґрунті, мулі, прісних та солоних водоймах, на тваринних і рослинних рештках, в організмі комах, тварин та людини. Є патогенні для людини види: B. anthracis - збудник сибірки; C. botulinum - збудник ботулізму; C. perfringens - збудник газової гангрени; C. tetani - збудник правцю. Деякі представники роду Bacillus, які вважались сапрофітами, віднесені до умовно-патогенних бактерій, наприклад B. cereus. Останніми роками описано ряд захворювань, які провокуються B. subtilis.
Група 19. Грампозитивні неспороутворювальні палички правильної форми. Бактерії, віднесені до даної групи, мають паличкоподібну форму, яка може варіювати від коротких паличок (майже коків) до довгих паличок, ниток або трихом, але всі вони мають правильну форму - плеоморфізм виражений слабо. Клітини грампозитивні, спор не утворюють, рідко пігментовані. Організми цієї групи - мезофіли, факультативні або облігатні анаероби, які здійснюють гомо- або гетероферментативне молочнокисле бродіння, здатні рости лише на складних поживних середовищах. Зустрічаються, як правило, в асоціаціях із рослинами чи тваринами або в органічній речовині, що розкладається. Є патогенні організми. Особливий інтерес становлять представники роду Lactobacillus і Listeria. Бактерії роду Lactobacillus виявляються в молоці, змивах молокопереробного обладнання, зернових і м'ясних продуктах, вині, фруктових соках, ротовій порожнині, кишковому тракті людини. Клітини - від довгих до коротких паличок (кокобацил), часто утворюють ланцюжки. Нерухомі, якщо рухомі, то мають перит- рихальний тип джгутикування. Грампозитивні, але при старінні культури і підвищенні кислотності середовища за Грамом не забарвлюються. У процесі зброджування цукрів утворюють лактат (до 50 %), ацетат, форміат, сукцінат, етанол та СО2. Типовим видом роду є L. delbrueckii.
Типовим видом роду Listeria є L. monocytogenes - збудник лістеріозу, гострого інфекційного захворювання, яке супроводжується ураженням заглоткових та інших залоз, мононуклеарною реакцією білої крові, часто - явищами септицемії.
Група 20. Грампозитивні неспороутворювальні палички неправильної форми. Більшість представників даної групи - це грампозитивні палички (Acetobacterium, Actinomyces, Arthrobacter, Bifidobacterium, Brevibacterium, Corynebacterium, Propionibacterium) неправильної форми, аероби (Arthrobacter, Brevibacterium), деякі рухомі. Інші - факультативні анаероби (Actinomyces, Corynebacterium, Propionibacterium), облігатні анаероби (Acetobacterium, Acetogenium, Bifidobacterium) або грамнегативні (Acetogenium).
Група 21. Мікобактерії. До цієї групи віднесено рід Mycobacterium. Клітини паличкоподібної форми, кислотостійкі, аеробні, ростуть повільно, вільноживучі або паразити хребетних. Кислотостійкість зумовлена наявністю восків у складі клітинної стінки, що особливо важливо для визначення мікобактерій. Клітини мікобактерій можуть утворювати нитки, які нагадують міцелій, але вони легко розпадаються на палички або коки. За Грамом забарвлюються з утрудненням, але вважається, що вони грампозитивні. Нерухомі, аероби і хемоорганотрофи. Типовий вид - M. tuberculosis.
Мікобактерії здатні до утворення спор. При цьому плазма розпадається на окремі ділянки (фрагменти), які стискаються й ущільнюються (рис. 7.10). Такі фрагменти і є самостійними зародковими тільцями - спорами. На утворення спор витрачається весь вміст клітини. При формуванні спор оболонка клітини ослизнюється, стає менш помітною, майже зникає, а спори вивільнюються. За способом утворення і біологічним значенням спори мікобактерій суттєво відрізняються від таких в ендоспороутворювальних бактерій. У клітині мікобактерій утворюються не одна, а декілька нестійких до дії несприятливих факторів спор.
Рис. 7.10. Спороутворення в мікобактерій:
а - концентрація плазми на окремі фрагменти; б, в - ущільнення фрагментів та їх перетворення у спори; г - розчинення оболонки клітин; д - зрілі спори
Групи 22-29. Актиноміцети. Грампозитивні бактерії, які здатні до утворення ниток (галуження) діаметром 0,5-1,0 мкм. Нитки можуть розпадатися на елементи різних розмірів або зберігатися цілими й утворювати артроспори. Спори поодинокі, у ланцюжках різної довжини або в спорангіях, як правило, нерухомі, але в деяких родів спори мають джгутики. Роди актиноміцетів відрізняються морфологічними ознаками, наприклад формою спороносців, а також наявністю маркерних хімічних компонентів клітинної стінки, мембран і гідролізатів цілих клітин (рис. 7.11). У більшості випадків це аероби, але деякі роди представлені факультативними чи облігатними анаеробами. Хемогетеротрофи. Деякі види патогенні для людини, тварин і рослин.

Рис. 7.11. Деякі типи спороносців актиноміцетів
Нокардіоформні актиноміцети утворюють нитки, які розпадаються на короткі елементи. Деякі роди утворюють повітряний міцелій з ланцюжками спор. Поділ на роди базується, перш за все, на хемотипі клітинної стінки, наявності чи відсутності міколових кислот та інших хімічних характеристиках. Ця підгрупа представлена родами Nocardia і Rhodococcus. Вегетативні гіфи роду Nocardia від рудиментарних до сильно розгалужених, діаметром 0,5-1,2 мкм, які ростуть на поверхні і проникають у товщу агаризованого середовища. Гіфи часто розпадаються на бактероїдні елементи - від паличко- до коко- подібних. Майже завжди утворюють повітряні гіфи. На повітряних, а інколи на субстратних гіфах виявляються ланцюжки спор. За Грамом забарвлюються позитивно або варіабельно. Аероби, частково кислотостійкі. Колонії мають плівчасту або бархатисту поверхню, коричневого, рижувато-коричневого, рожевого, оранжевого, червоного, пурпурового, сірого або білого кольору, гладенькі або зернисті. У представників роду Rhodococcus можливе утворення різних морфологічних форм - від паличок до розгалуженого вегетативного міцелію. Морфогенетичний цикл розпочинається зі стадії коків або коротких паличок, які переходять у послідовний ряд морфологічних змін, що завершують життєвий цикл. Так, коки можуть проростати в короткі палички, утворювати нитки з бічними виростами чи елементами галуження або сильно розгалужені гіфи. Наступна генерація коків або коротких паличок утворюється під час фрагментації паличок, ниток та гіфів. Грампозитивні, частково кислотостійкі, аероби.
До родів із багатогніздовими спорангіями віднесено: Dermatophilus, Frankia, Geodermatophilus. Повітряний міцелій у представників роду Dermatophilus розвивається в атмосфері СО2. Субстратний міцелій складається з довгих, звужених на кінці ниток, які галузяться латерально під прямими кутами. Перегородки, які утворюються в поперечній, горизонтальній і вертикальній площинах дають початок восьми паралельним рядам кокоподібних клітин (спор), кожна з яких рухається за допомогою джгутика. Грампозитивні, аероби і факультативні анаероби, які ростуть на складних поживних середовищах. Паразити ссавців (переважно сільськогосподарських тварин). D. congolensis є збудником ексудативного дерматиту. Представники роду Frankia. не утворюють повітряного міцелію. Вегетативні гіфи слабко або сильно галузяться. Багатогніздові спорангії, округлі або неправильної форми, утворюються термінально, латерально чи інтеркалярно на вегетативних гіфах. Спорангіоспори нерухомі, розміром 1,0-5,0 мкм, від незабарвлених до чорних. Міколати не виявляються. Аероби або мікроаерофіли. Ростуть повільно (час генерації 17 діб). Більшість штамів здатні фіксувати атмосферний азот in vitro і в рослинах - вони є симбіонтами ряду покритонасінних рослин, які індукують утворення бульбочок (F. alni).
Актиноплани утворюють тонкий збіднений міцелій, який галузиться, але не розпадається. Утворюють пігменти різних кольорів, які добре дифундують у середовище. Спори сферичної форми утворюються всередині спорангіїв (спорових везикул), розташовані на спорангіофорах або сидячі. Потрапляючи у воду, спори вивільнюються із спорангіїв, рухаються за рахунок полярних джгутиків. Грампозитивні, але частина гіф може не забарвлюватися за Грамом. Виявляються в ґрунті та рослинному матеріалі, який розкладається, - Actinoplanes philippinensis. Представники роду Micromonospora мають добре розвинений, розгалужений і септований міцелій діаметром 0,5 мкм. Нерухомі спори утворюються по одній на коротких або довгих спороносцях, які часто розташовані розгалуженими пучками (рис. 7.12). Спороносці утворюються моноподіально чи симподіально. Повітряний міцелій не виявляється. Грампозитивні, не кислотостійкі, хемоорганотрофи. Виявляються в ґрунтах, воді, морському середовищі, мулі - M. chalcea.
Рис. 7.12. Утворення спор у мікромоноспор:
а - Micromonospora - спори сидять на поверхні у вигляді горбочків або коротких ланцюжків на коротких ніжках-спороносцях; б - Microbispora - двоклітинні спори на коротких ніжках; в - Micropolyspora - ланцюжки спор, прямі або спірально зігнуті; г - Actinobifida - дихотомічне галуження пороносних гілок, по одній спорі на кінці гілки
Стрептоміцети і близькі роди. Актиноміцети цієї групи утворюють міцелій, який галузиться (діаметром близько 1,0 мкм) і має тенденцію розпадатися на фрагменти. Рід Streptomyces має вегетативні гіфи діаметром 0,5-2,0 мкм, які утворюють сильно розгалужений міцелій, що рідко розпадається на фрагменти (S. albus, S. antibioticus, S. diastaticus,
S. albidoflavus, S. gHseoviHdis). Повітряний міцелій у зрілому стані несе ланцюжки з трьох чи більше нерухомих спор. Деякі види можуть формувати склероціо-, пікнідіо- і спорангіоподібні структури. Колонії шкірясті або маслянисті. Поверхня колонії на початку гладенька, але з часом формується переплетення повітряного міцелію, яке може бути пластів - цевого, зернистого, порошкоподібного чи бархатистого типу. Утворюють різні пігменти, які визначають колір субстратного і повітряного міцелію. Часто синтезують також пігменти, які дифундують у середовище. Багато штамів продукують один або декілька антибіотиків. Оптимум температури - 25-35 0С, оптимальний для росту діапазон pH - 6,5-8,0. Значно поширені в ґрунті, включаючи компости. Деякі види патогенні для тварин і людини, інші - фітопатогени. Рід Streptoverticillium має розгалужений субстратний міцелій діаметром 0,8-1,2 мкм. Повітряний міцелій складається з довгих прямих гіфів, на яких розташовані мутовки гілочок. Кожна гілочка-мутовка має на кінчику зонтик з ланцюжка сферичних та еліпсоподібних шипуватих чи волосистих спор (S. baldaccii, S. griseocameum, S. viridoflavum).
Група 30. Мікоплазми (молікути): бактерії без клітинної стінки. Мікоплазми не мають істинної клітинної стінки, не здатні синтезувати таких її попередників, як мурамова і диамінопімелінова кислоти. Клітини дуже дрібні, інколи до ультрамікроскопічних (~200 нм), плеоморфні. За морфологією клітин - від коків до ниток, які здатні до утворення міцелієподібних структур. Розмноження відбувається за рахунок утворення в нитках кокоподібних структур - елементарних тілець, визначається також бінарний поділ і брунькування. За Грамом не забарвлюються.
Мікоплазми (відділ Tenericutes) об'єднані у клас Mollicutes, який має порядок Mycoplasmatales з трьома родинами: Mycoplasmataceae, Acholeplasmataceae і Spiroplasmataceae. Поділ мікоплазм на родини базується на їх вимогах до стеринів, стійкості до дигітоніну, наявності лактатдегідрогенази, величині геному, здатності до синтезу жирних кислот з ацетату тощо. Так, стеринозалежні мікоплазми родини Mycoplasmataceae потребують стеринів, які є основним компонентом мембранних ліпідів паразитичних форм. Стеринозалежні мікоплазми родини Spiroplasmataceae із спіралеподібною формою клітини характеризуються в деяких фазах (у lag-фазі - сферичні й короткі спіральні клітини, у стаціонарній - довгі) здатністю до обертального і гвинтоподібного руху (джгутики чи аксіальні фібрили відсутні).
Родина Mycoplasmataceae представлена двома родами: Mycoplasma і Ureaplasma. Діаметр кулястої клітини коливається від 125 до 250 нм, а довжина ниткоподібних форм може досягати 150 мкм. При старінні культури нитки розпадаються на ланцюжки кокоподібних клітин. На щільних поживних середовищах вони утворюють дрібні, напівсферич- ні, сплющені колонії діаметром 10-600 мкм, інколи колонія досягає 4 мм. Типові колонії двофазні і мають вигляд "вилитої яєчні». Вони складаються з каламутної, гранулярної центральної частини, яка вростає в середовище, і плоскої периферійної зони, що просвічується. Розмножуються бінарним поділом, брунькуванням або множинним вивільненням елементарних тілець. Елементарні тільця - це структури ~ 100 нм, які виявляються в загальній масі колонії. Вважається, що вони утворюються внаслідок якихось дегенеративних процесів, які мають місце в екстремальних умовах, тобто в умовах, коли клітина має загинути. При загибелі клітини вивільнюються частинки типу спор, які, потрапивши в оптимальні умови, дають початок новому поколінню. Мікоплазми є паразитами широкого кола хазяїв - ссавців і птахів. Вони можуть викликати гострі і хронічні патологічні процеси, які перебігають за типом латентних інфекцій. Доведена роль у патології людини M. pneumoniae. M. hominis часто висівається з урогенітального тракту і, можливо, провокує уретрити та інші запальні процеси. M. arthritidis висівається із синовіальної рідини уражених суглобів. M. salivarium і M. oral є коменсалами ротової порожнини. Типовим видом роду є M. agalactiae. Особливістю представників роду Ureaplasma (U. diversum, U. felinum, U. urealiticum) є здатність гідролізувати сечовину.
Ахолеплазми не потребують для свого розвитку стеринів. На щільних поживних середовищах вони утворюють досить великі колонії (~ 3 мм). Це факультативні анаероби, хемоорганотрофи з бродильним типом метаболізму. Серед них є вільноживучі сапрофіти, паразити і, можливо, патогени ссавців і птахів. Типовий вид - Acholeplasma laidlawii.
Опис родини Spiroplasmataceae вперше зробив І. Скрипаль у 1978 р. Діаметр сферичної клітини коливається від 0,1 до 0,25 мкм. У рідкому середовищі домінують спіральні клітини діаметром ~ 0,12 мкм і завдовжки до 2-4 мкм. На щільних середовищах формуються округлі, неспіралеподібні форми. На щільних субстратах утворюють характерні для молікутів колонії - типу яєчні з чіткими краями. На середовищі з вологою поверхнею формуються гранульовані колонії з дифузною межею. По краях колонії з'являються "дочірні" колонії, які утворюються в результаті пересування окремих клітин з первинної колонії. Розмір колонії коливається в межах 0,2 мм. Факультативні анаероби. Температурний діапазон - 20-37 0С, температурний оптимум - 32 0С. Виділяються з листків цитрусових рослин, але захворювання не відтворюється експериментально. Типовий вид - Spiroplasma citri.
За філогенетичною систематикою відділ Tenericutes скасовано, а клас Mollicutes входить до групи грампозитивних бактерій (Firmicutes). Філогенентични зв'язки класу молікут а також шляхи їхньої еволюції дискутуються до цього часу. Так, на базі порівняльного аналізу 16S рРНК мікоплазм із грампозитивними бактеріями було підтверджено їхню спорідненість з представниками роду Clostridium.
Група 31. Метаногени. Грамнегативні або грампозитивні бактерії, у яких клітинна стінка не має ні муреїну, ні зовнішньої мембрани. Мембрани складаються з ізопреноїдів, зв'язаних ефірними зв'язками.
Облігатні анаероби. Хемоавтотрофи або хемогетеротрофи, які завжди утворюють метан як продукт метаболізму. Крім метаногенів, жодні інші організми метан не утворюють. Як джерело вуглецю й енергії використовують H2 + CO2, форміат, ацетат, сполуки, які містять метильну групу (метанол, метиламіни, метилсульфіди) та ін. Клітина може бути паличко-, ланцето- або кокоподібної форми; асимілюють H2 + CO2, форміат або Н2 + метанол. Клітинна стінка містить псевдомуреїн - MethanobacteHum (M. formicicum), Methanobrevibacter (M. ruminantium), Methanosphaera (M. stadtmanae), Methanothermus (M. fervidus). Бактерії, які мають клітини коко-, паличко-, спірале- або пластинкоподібної форми, використовують Н2 + СО2, форміат або спирти + СО2, а клітинні стінки не містять псевдомуреїну, віднесено до родів: Methanococcus (M. vannielii), Methanogenium (M. cariaci), Methanomicrobium (M. mobile), Methanospinllum (M. hungatei). Псевдосарцини, коки або покриті чохлом палички, які використовують як субстрат для росту триметиламін або ацетат, становлять підгрупу, яка об'єднує роди: Methanococcoides (M. methylutens), Methanolobus (M. tindarius), Methanosarcina (M. barkeri).
Група 32. Сульфатредукуючі археї. Мають клітини неправильної форми, часто трикутні, діаметром 0,4-1,3 мкм, поодинокі, у парах, із джгутиками або без них. За Грамом не забарвлюються. При освітленні світлом із довжиною хвилі 420 нм дають блакитно-зелену флуоресценцію. На щільних середовищах утворюють зеленкувато-чорні гладенькі колонії діаметром 1-2 мм. Облігатні анаероби. Хемолітотрофи, хемоор- ганотрофи або хемоміксотрофи. Для автотрофного росту потребують тіосульфат і Н2, у присутності сульфату ріст слабкий. Утворюють H2S. Можуть відновлювати S0. Температурний діапазон росту 60-95 0С, температурний оптимум - близько 83 0С. Діапазон pH - 4,5-7,5, оптимум - 6,0. Виділяються з мілководних і глибоководних морських гідротермальних екосистем. До даної групи віднесено рід Archaeoglobus (A. fulgidus).
Група 33. Екстремально галофільні аеробні архебактерії (гало-бактерії). Представлені організмами кокоподібної форми (0,82,0 мкм) або неправильних паличок (0,3-1,2 х 1,0-15,0 мкм). Рухомі за рахунок пучків полярних джгутиків або нерухомі. Грамнегативні (палички) або грамваріабельні (коки). Паличкоподібні клітини лізуються при суспендуванні в дистильованій воді. Колонії забарвлені в червоний колір різних відтінків за рахунок вмісту у клітинах каротиноїд- них пігментів. Аероби, але деякі представники здатні до анаеробного росту у присутності нітрату. Хемогетеротрофи, потребують для росту не менш як 1,5М NaCl; для більшості оптимальна концентрація NaCl - 2,0-4,0 М. Виявляються в солоних і лужних озерах, солеварнях та засолених ґрунтах: Halobacterium (H. salinarium), Halococcus (H. morrhuae), Natronobacterium (N. gregoryi), Natronococcus (N. occultus).
Група 34. Архебактерії, позбавлені клітинної стінки. Група представлена родом Thermoplasma, який об'єднує плеоморфні клітини з варіаціями форми від сферичної (0,1-5,0 мкм) до ниткоподібної. Клітини позбавлені клітинної стінки й оточені лише клітинною мембраною завтовшки близько 7 нм. Ліпіди клітинної мембрани представлені дибіфітанильними (С40) дигліцерольними тетраефірами. Грамнегативні, можуть бути рухомими за рахунок джгутиків. Облігатні термофіли (діапазон температур для росту 33-67 0С); облігатні ацидофіли (ростуть при pH 0,5-4,0). За умов нейтральних pH відбувається лізис клітини. Факультативні анаероби. Анаеробний ріст стимулюється елементарною сіркою, яка відновлюється до H2S. Поширені у вугільних териконах. Типовий вид - T. acidophilum.
Група 35. Екстремальні термофіли і гіпертермофіли, які метаболізують S0. Це грамнегативні рухомі або нерухомі палички, нитки, коки або клітини дископодібної форми. Серед них є аероби, факультативні або облігатні анаероби. Ростуть хемоавтотрофно або хемогетеротрофно. В анаеробних умовах окиснюють H2S або S0до H2SO4. Донорами електронів служать Н2 або органічні речовини. Діапазон температур для росту - 45-110 0С, оптимальна температура - 70-105 0С. Поширення - континентальні сольфатарні поля або морські гідротермальні системи. Група розділена на три підгрупи. Підгрупа 1 об'єднує термоацидофільні аеробні або факультативно анаеробні кокоподібні організми, які ростуть при pH нижче 4,0: рід Acidianus (A. infernus, температурний оптимум - 88 0С, максимальна температура для росту - 96 0С; A. brierleyi, оптимальна температура для росту - 70 0С, максимальна - 75 0С); рід Sulfolobus (S. acidocaldarius, діапазон температур - 55-87 0С, діапазон pH - 1,0-6,0). До підгрупи 2 належать паличко- або ниткоподібні бактерії, облігатні анаероби, які ростуть при pH понад 4,0. Рід Thermofilum (T. pendens, клітини паличкоподібної форми діаметром 0,15-0,35 мкм і завдовжки від 1,0 до ~100 мкм; діапазон pH - 4,0-6,7; інтервал температур - 70-95 0С). Підгрупа 3 об'єднує облігатні анаероби коко- або дископодібної форми, які ростуть при pH понад 4,0. Бактерії роду Desulfurococcusмають кокоподібні клітини грампозитивного типу, рухомі або нерухомі. Діапазон температури - 70-95 0С (оптимальна - 85 0С), діапазон pH - 4,57,0 (оптимальна - близько 6,0). Типовий вид - D. mucosus.