ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 20. Иммунология опухолей
ОПУХОЛЕАССОЦИИРОВАННЫЕ АНТИГЕНЫ, РАСПОЗНАВАЕМЫЕ КЛЕТКАМИ ИММУННОЙ СИСТЕМЫ
Существование опухолевых антигенов впервые было обнаружено при постановке трансплантационных тестов. Когда опухоль пересаживали животному, предварительно иммунизированному инактивированными клетками той же опухоли, трансплантат отторгался. Резистентность к пересаженной опухоли, опосредованная, как впоследствии было установлено, клетками иммунной системы, направлена па опухолеассоциированные трансплантационные антигены двух типов. Антигены первого типа (Т-, от англ. tumor, антигены) — общие для многих опухолей, даже различного тканевого происхождения. Антигены второго типа специфичны для каждой отдельной опухоли — это опухолеспецифические трансплантационные антигены. Возможна одновременная экспрессия специфических и Т-антигенов.
Т-антигены имеют вирусное происхождение
Эти антигены обнаруживаются на клетках опухолей, индуцированных вирусами, например мелким ДНК-содержащим вирусом полиомы и вирусом SV40 (эти два вируса способны вызывать опухоли у экспериментальных животных) и вирусами папилломы (они ассоциированы с раком шейки матки у человека). Данные вирусы кодируют Т-антигены, свойственные и другим вирусам той же группы. Эти антигены представляют собой ядерные белки, играющие определенную роль в поддержании трансформированного состояния.
Инфекционные РНК-содержащие онкогенные вирусы вызывают лейкозы и саркомы у животных; обнаружен также по крайней мере один вирус Т-клеточного лейкоза человека (HTLV-1) (рис. 20.3). Эти вирусы выходят из инфицированных клеток путем отпочковывания от их клеточной мембраны, приобретая при этом оболочку; в мембране инфицированных ими клеток выявляется гликопротеин вирусной оболочки. Общие антигены ДНК-, как и PHК-содержащих онкогенных вирусов вызывают сильный гуморальный и клеточный ответ, способный обеспечить защиту против опухоли. Поскольку опухоли, вызываемые данным онкогенным вирусом, экспрессируют один и тот же антиген, инбредные мыши, иммунизированные, например, многократными инъекциями облученных клеток опухоли, индуцированной SV40, отторгают опухоли, вызванные данным вирусом, но чувствительны к опухолям, вызываемым вирусом полиомы.
Для некоторых линий мышей характерна спонтанная активация эндогенных РНК-содержащих онкогенных вирусов, приводящая к развитию лейкоза. У животных других линий опухоли, которые могут экспрессировать вирусные антигены и продуцировать инфекционный вирус лейкоза мышей (MuLV), образуются в том случае, если им вводить канцерогенное химическое соединение. Такие опухоли экспрессируют как общие опухолеассоциированные антигены, так и опухолеспецифические антигены, рассматриваемые ниже. У организма-хозяина, однако, эндогенные РНК-содержащие вирусы вызывают лишь слабый иммунный ответ, возможно, из-за иммунологической толерантности (см. гл. 14).
Специфические опухолевые антигены отражают изменения в опухолевых генах или в экспрессии генов
К специфическим опухолевым антигенам относят те антигены, которые могут вызвать иммунный ответ на введенные опухолевые клетки, в том случае если животное было предварительно иммунизировано материалом той же опухоли (рис. 20.4). Такие антигены впервые были выявлены при изучении опухолей, индуцированных у инбредных мышей канцерогенными химическими веществами, и к настоящему времени их природа установлена в описываемых ниже экспериментах.
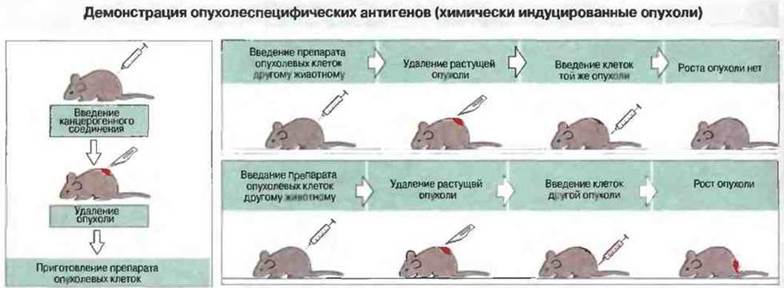
Рис. 20.4. У мышей индуцировали образование опухолей введением химического канцерогенного соединения (метилхолантрена). Затем опухолевые клетки этих животных подкожно иньецировали генетически идентичным мышам. Растущие опухоли удаляли хирургическим путем, после чего животным вводили клетки той же опухоли; в этом случае опухоль не развивалась. При введении клеток другой опухоли (индуцированной тем же канцерогеном) опухоль развивалась. Способность к отторжению опухоли может быть передана лимфоидными клетками.
Перевиваемую (т. е. слабоиммуногенную) культуру опухолевых клеток от мыши инбредной линии подвергали in vitro воздействию сильного мутагена. Затем из этой культуры получали субклоны мутантных опухолевых клеток, часть которых была неспособна к дальнейшему росту in vivo, если только имплантат не был очень массивным. Мутантные культуры обнаруживали большую иммуногенность, чем родительские опухолевые клетки. При иммунизации одним из этих опухолеотрицательных (tum-) клонов у генетически идентичных мышеи образовывались Тц-клетки, способные уничтожать имплантат только того клона, который был использован для иммунизации, а не родительской или других tum--культур (рис. 20.5). Эти Тц-клетки затем использовали в качестве зондов для определения мутантного опухолевого антигена при молекулярном клонировании соответствующего гена. В итоге мутантный ген, кодирующий данный опухолевый антиген (tum--ген), был идентифицирован и секвенирован. Сравнение этого гена с гомологичным геном из клеток родительской опухоли показало различие их продуктов всего по одному аминокислотному остатку. В дальнейшем было четко доказано, что такая мутация приводит к образованию иммуногенного антигена, распознаваемого Тц-клетками: если клетки родительском опухоли инкубировали с пептидом из 10 аминокислотных остатков, содержащим tum--последовательность, эти клетки затем уничтожались Тц-клетками, однако в случае инкубации с гомологичным пептидом из клеток родительской опухоли лизиса не происходило (рис. 20.6). Учитывая, что распознавание антигенов Tц-клетками рестриктировано по молекулам МНС класса I, можно предположить, что опухолеспецифичный белок процессируется в опухолевой клетке с образованием пептида, который затем образует комплекс с антигенами МНС класса I и транспортируется на клеточную поверхность.
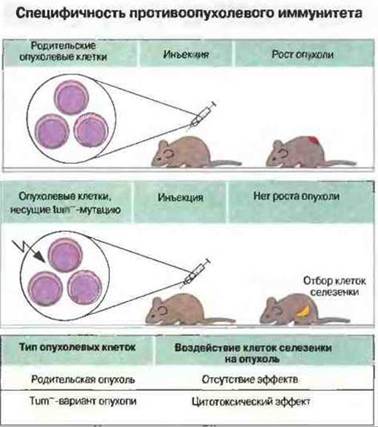
Рис. 20.5. Образование высокоиммуногенного клона опухолевых клеток (tum-) мышей DBA2 и специфичность Тц-клеток к данной опухоли. После индуцирования мутаций в клетках родительской опухоли из них были получены субклоны, часть которых оказалась неспособной к росту in vivo при введении мышам DBA2. Тц-клетки из селезенки животных, которым были введены tum--клетки, уничтожали опухолевые клетки tum--клона, но не родительской линии.
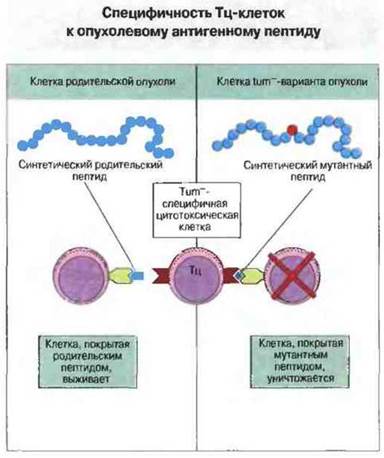
Рис. 20.6. Тц-клетки мышей, иммунизированных опухолевыми клетками tum--клона, in vitro уничтожают опухолевые клетки, покрытые пептидом — продуктом tum--гена, но не клетки, покрытые гомологичным пептидом из родительской опухоли. Два этих пептида различаются одним аминокислотным остатком.
Клонированные гены опухолевых антигенов из клеток других tum--клонов иногда оказывались идентичными родительскому гену. Отличие таких tum--клонов от родительской линии заключалось в том, что в tum--клетках происходила сверхэкспрессия данного антигена. Получены убедительные доказательства в пользу существования иммунного ответа, рестриктированного по антигенам МНС класса II, по крайней мере на опухоли человека, однако значительно меньше известно о том, какие опухолевые антигены распознаются в комплексе с антигенами МНС класса II.