ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 26. Гиперчувствительность - тип IV
■ Известны три реакции гиперчувствительности IV типа: контактная, туберкулиновая и гранулематозная.
■ Нанесенный на кожу гаптен поглощают и процессируют клетки Лангерганса; эти же клетки презентируют гаптен антигенспецифичным Т-клеткам.
■ Цитокины, продуцируемые иммунокомпетентными клетками кожи (например, кератиноцитами, клетками Лангерганса или Т-клетками), обусловливают участие в реакции антиген- неспецифичных Т-клеток и макрофагов.
■ Реакцию гиперчувствительности туберкулинового типа можно использовать в качестве диагностической пробы на присутствие многих инфекционных агентов.
■ Для гранулематозных реакций характерен определенный баланс между защитным иммунитетом к нерастворимому антигену и опосредованным Т-клетками повреждением ткани. Характерный пример такого равновесия - туберкулоидная форма проказы.
■ Персистенция антигена приводит к дифференцировке макрофагов в эпителиоидные клетки и их слиянию с образованием гигантских клеток. Этот патологический процесс, называемый гранулематозной реакцией, приводит к повреждению тканей.
■ Формирование гранулемы связано с Т-клеточной активацией макрофагов и зависит от ФНО.
Согласно классификации Кумбса и Джелла, гиперчувствительность IV типа (замедленная) — это реакции, проявляющиеся не ранее, чем через 12 ч и опосредованные клеточными, а не гуморальными иммунными механизмами. Однако для некоторых реакций гиперчувствительности такая классификация не подходит. Например, реакции поздней фазы, достигающие пика через 12-24 ч после контакта с аллергеном, опосредованы преимущественно IgE, хотя в них принимают участие и Т-хелперные клетки, т. е. механизм их комплексный. Другие реакции (например гиперчувствительность Джонса-Моута, которая напоминает кожные базофильные реакции у морских свинок) раньше также относили к IV типу, хотя их механизмы и клиническое значение остаются неясными; в этой главе они не рассматриваются.
В отличие от других видов гиперчувствительности, реакции IV типа могут быть перенесены от сенсибилизированного животного несенсибилизированному не сывороткой, а Т-клетками (у мыши Тц1-клетками). В основе таких реакций лежат несомненно механизмы защитного Т-клеточного иммунитета, однако полная корреляция между ним и гиперчувствительностью IV типа наблюдается далеко не всегда. Ответственные за осуществление замедленной реакции Т-клетки специфически сенсибилизированы антигеном в период предшествующего контакта с ним и действуют путем привлечения к месту реакции клеток других типов.
Известны три варианта реакций гиперчувствительности IV типа (рис. 26.1). Контактная и туберкулиновая реакции развиваются в течение 72 ч после начала действия антигена, тогда как гранулематозная реакция — лишь через 21—28 сут; гранулемы образуются в результате накопления и пролиферации макрофагов и могут сохраняться неделями. Эта форма гиперчувствительности IV типа вызывает наиболее серьезные клинические последствия. Необходимо отметить, что один и тот же антиген способен вызывать реакции разных видов, которые могут перекрывать друг друга.
Названные три вида реакций замедленного типа исходно были выделены по характеру проявлений, возникающих при накожном нанесении или внутрикожном введении антигена. Степень реакции обычно оценивают у животных по толщине пораженного участка кожи. Вслед за местной реакцией могут развиваться и различные системные иммунные реакции.
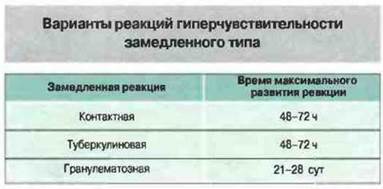
Рис. 26.1. Контактная и туберкулиновая формы гиперчувствительности имеют сходную временную динамику и достигают максимума через 48-72 ч. В некоторых случаях (например, при воздействии нерастворимого антигена) через 21-28 сут развивается и гранулематозная реакция (например, при кожной пробе на проказу).
КОНТАКТНАЯ ГИПЕРЧУВСТВИТЕЛЬНОСТЬ
Контактная гиперчувствительность характеризуется экзематозной реакцией в месте воздействия антигена (рис. 26.2). Она часто возникает в результате контакта с такими веществами, как никель, хромат, применяемые в резиновой промышленности катализаторы и пентадекакатехол (присутствующий в растении сумах укореняющийся). При контакте с токсически действующими раздражающими веществами экзема может возникать и без участия механизмов гиперчувствительности. Хотя начальные реакции в этих двух случаях различны, иммунологические сдвиги, развивающиеся после воздействия раздражителей и аллергенов, сходны между собой.
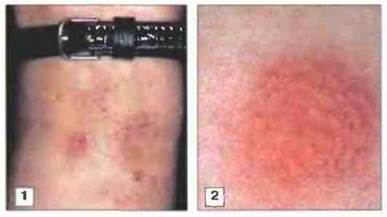
Рис. 26.2. Клинические проявления и результат кожной пробы при контактной реакции гиперчувствительности. 1. Экзематозные изменения на запястье, вызванные никелевой застежкой от часов. 2. Подозрение на аллергию можно подтвердить, нанося потенциальные аллергены (в достаточной концентрации и в комплексе с подходящим носителем) на кожу внутренней поверхности предплечья (кожная проба). При положительной реакции через 2-4 сут на месте нанесения вызывающего аллергию антигена появляется ограниченный участок экземы.
Иммунологически активные компоненты перечисленных выше агентов названы гаптенами. Сами по себе гаптены не обладают антигенными свойствами (размеры их молекул для этого слишком малы — молекулярная масса часто не достигает и 1 кДа), но, проникая в эпидермис и присоединяясь (чаще всего ковалентно) к собственным белкам организма, они приобретают антигенную активность. Сенсибилизирующую активность гаптенов трудно предсказать на основании их структуры, хотя имеется некоторая корреляция между количеством связанного с носителем гаптена и его способностью проникать в кожу. Отдельные контактные аллергены содержат ненасыщенные углеродные связи и легко окисляются. Некоторые гаптены, например динитрохлорбензол (ДНХБ), сенсибилизируют почти любого человека и могут быть использованы для оценки клеточного иммунитета. Нанесенный на кожу ДНХБ связывается с белками эпидермиса через NH2-группы остатков лизина.
Ключевая роль при контактной гиперчувствительности принадлежит клеткам Лангерганса и кератиноцитам
Клетки Лангерганса служат основными антигенпрезентирующими клетками. Контактная гиперчувствительность - это главным образом эпидермальная реакция, и основными антигенпрезентирующими клетками (АПК) в данном случае служат дендритные клетки Лангерганса, локализованные в надбазальном слое эпидермиса (рис. 26.3). Эти клетки имеют костномозговое происхождение и экспрессируют на своей поверхности маркер CDI, антигены МНС класса II, а также рецепторы для Fc и комплемента (см. гл. 2). Как показывает электронная микроскопия, клетки Лангерганса содержат бербековы гранулы (специфические для этих клеток органеллы, образующиеся из клеточной мембраны). Под действием В-ультрафиолета клетки Лангерганса инактивируются, что предотвращает или смягчает проявления контактной гиперчувствительности.
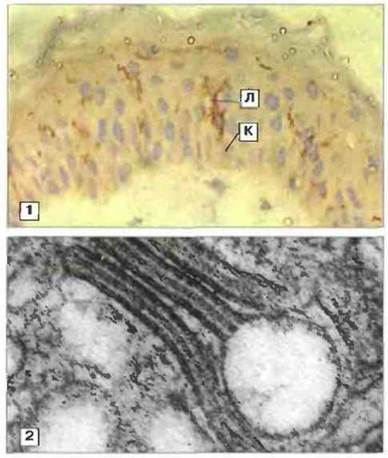
Рис. 26.3. Клетка Лангерганса.
1. Эти дендритные клетки составляют 3 % всех клеток эпидермиса. Они экспрессируют ряд поверхностных маркеров, по которым их можно визуализировать. В данном случае срез нормальной кожи окрашен моноклональными антителами к CD1 (второй краситель - гемалум Майера).
(Л - клетка Лангерганса; К - кератиноцит.) х 312.
2. На электронной микрофотографии клетки Лангерганса видна характерная бербекова гранула. Эта пластинковидная органелла имеет четкую центральную исчерченность и часто расширена с одного конца. х 132 000.
In vitro клетки Лангерганса действуют как АПК, будучи в этом отношении более активными, чем моноциты. Однако механизм процессинга антигена в клетках Лангерганса остается неясным. Антигены, связанные с молекулами МНС класса II, могут поглощаться ими путем опосредованного рецепторами эндоцитоза с участием бербековых гранул.
Кератиноциты продуцируют ряд важных для реакций контактной гиперчувствительности цитокинов
Кератиноциты обеспечивают структурную целостность эпидермиса и выполняют центральную роль в иммунных реакциях этого слоя кожи. Они могут экспрессировать на мембране молекулы МНС класса II и ICAM-1, а также выделять цитокины, в том числе ИЛ-1, ИЛ-3, ИЛ-6, ИЛ-8, ГМ-КСФ, ФНОα, ТФРα и ТФРβ. Интерлейкин-3 способен активировать клетки Лангерганса, участвовать в стимуляции пролиферативного ответа и привлекать тучные клетки, а также индуцировать секрецию иммуносупрессивных цитокинов (например, ИЛ-10 и ТФРβ), которые подавляют иммунный ответ и вызывают клональную анергию (иммунологическую ареактивность) Тх1-клеток.
Кератиноциты активируются многими стимулами, включая аллергены и раздражающие вещества, и в активированном состоянии продуцируют иммунорегуляторные цитокины, такие как ФНОα и ГМ-ГСФ, которые в свою очередь активируют клетки Лангерганса. Некоторые антигены, в том числе урушиол из сумаха укореняющегося, непосредственно индуцируют образование ФНОα и ИЛ-8.
Реакция контактной гиперчувствительности протекает в две стадии: сенсибилизации и проявления
В процессе сенсибилизации образуются Т-клетки иммунологической памяти. У человека период сенсибилизации длится 10—14 сут. Гаптен, проникнув в кожу, соединяется с белком, после чего происходит его поглощение (интернализация) эпидермальными клетками Лангерганса. Эти клетки затем покидают эпидермис и по афферентным лимфатическим сосудам мигрируют в паракортикальную область регионарных лимфоузлов. Здесь они презентируют подвергшиеся процессингу гаптен-белковые конъюгаты (ассоциированные с молекулами МНС класса II) лимфоцитам CD4+, формирующим популяцию клеток иммунологической памяти (рис. 26.4). Исследования на человеке с применением ДНХБ показывают, что сенсибилизация зависит прежде всего от дозы гаптена на единицу площади кожи, а не от обшей дозы или общей площади, на которую воздействует гаптен.
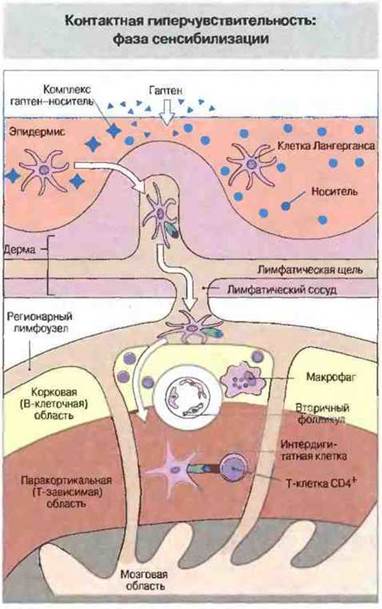
Рис. 26.4. В эпидермисе образуется комплекс гаптена с белком-носителем. Клетки Лангерганса поглощают (интернализуют) антиген и по афферентным лимфатическим сосудам мигрируют в паракортикальную область регионарного лимфоузла, где в качестве интердигитатных клеток презентируют антиген Т-клеткам CD4+.
Фаза проявления реакции протекает с участием лимфоцитов CD4+и моноцитов
В течение нескольких часов после нанесения аллергена на кожу обычно наблюдается некоторое снижение количества клеток Лангерганса в эпидермисе. Затем в коже и лимфоузлах эти клетки презентируют антиген. Вскоре происходит дегрануляция тучных клеток с выделением цитокинов. Продуцируемые клетками разных типов, в частности макрофагами, цитокины ФНОα и ИЛ-1 служат мощными индукторами выделения молекул адгезии эндотелиальными клетками. В результате местного выделения этих цитокинов формируется градиент их концентрации, что служит сигналом для движения мононуклеарных клеток в направлении дермо-эпидермального контакта и эпидермиса. Фазу проявления контактной гиперчувствительности иллюстрирует рис. 26.5.
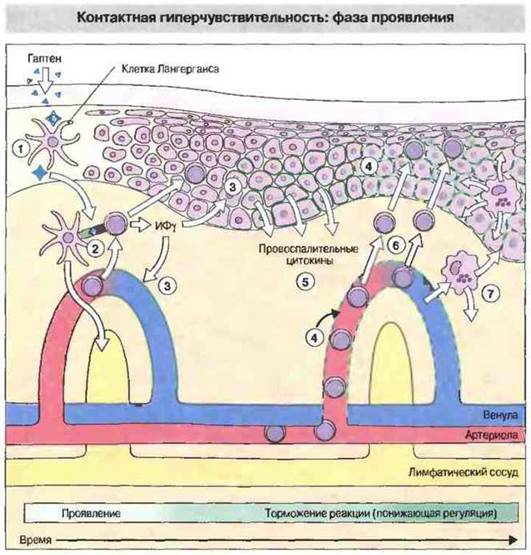
Рис. 26.5. Клетки Лангерганса, несущие комплекс гаптен-носитель (1) перемещаются из эпидермиса в дерму, где презентируют этот комплекс Т-клеткам (CD4+) иммунологической памяти (2). Активированные Т-клетки CD4+выделяют ИФγ, который индуцирует экспрессию 1САМ-1 (3) и, позднее, молекул МНС класса II (4) на поверхности кератиноцитов и на эндотелиальных клетках кожных капилляров, а также стимулирует кератиноциты, вызывая продукцию ими провоспалительных цитокинов, таких как ИЛ-1, ИЛ-6 и ГМ-ГСФ (5). Цитокины привлекают к месту реакции антигеннеспецифические Т-клетки CD4+(6), которые могут связываться с кератиноцитами через ICAM-1 и молекулы МНС класса II. К очагу реакции привлекаются также активированные макрофаги, но это происходит позднее. С данного момента реакция начинает угасать. В супрессии реакции могут участвовать эйкоэаноиды, например ПГЕ, выделяемый активированными кератиноцитами и макрофагами (7).
Наиболее раннее (через 4—8 ч) гистологическое изменение состоит в появлении мононуклеарных клеток вокруг производных кожи (ногти, волосы) и кровеносных сосудов; в дальнейшем они инфильтрируют эпидермис. Макрофаги проникают в дерму и эпидермис через 48 ч. Количество инфильтрирующих эти слои кожи клеток достигает максимума через 48—72 ч (рис. 26.6), причем большинство лимфоцитов относится к популяции CD4+и лишь немногие к CD8+. Менее 1% инфильтрирующих кожу клеток принадлежит к одному клону (а именно к клону антигенспецифичных клеток иммунологической памяти — Тх1-клеток CD4+). Привлечение Т-клеток происходит в основном по независимому от антигена механизму.
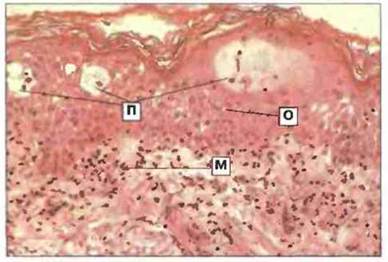
Рис. 26.6. Гистологическая картина повреждения при контактной реакции гиперчувствительности. Мононуклеарные клетки (М) инфильтрируют дерму и эпидермис. Эпидермис набухает и в нем вследствие отека (О) появляются микропузырьки (П). Окраска гематоксилином и эозином, х 130.
Механизмы реакций на аллергены и на раздражающие вещества частично сходны
И аллергены, и раздражающие вещества могут повреждать клетки Лангерганса и индуцировать выделение цитокинов, способствующих созреванию этих клеток в активные АПК, которые мигрируют в лимфоузлы и инициируют иммунный ответ. Синтез мРНК для ФНОα, ИФγ и ГМ-КСФ начинается уже в первые 30 мин после нанесения на кожу аллергена или раздражающего вещества, а через 2—4 ч содержание этих мРНК возрастает в 10 раз. Некоторые изменения в транскрипции мPHК отмечаются только после воздействия гаптена. К ним относится увеличение уровня мРНК для ИЛ-1β в клетках Лангерганса (через 15 мин), а также усиление транскрипции мРНК для ИЛ-1α в кератиноцитах, воспалительного белка-2 макрофагов (М1Р-2) и индуцируемого интерфероном белка-10 (IP-10) (рис. 26.7).
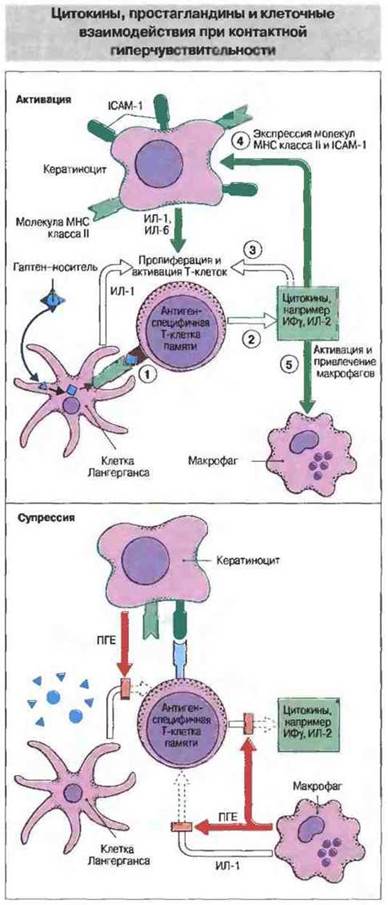
Рис. 26.7. Центральная роль в сложных взаимодействиях между клетками Лангерганса, Т-клетками CD4+, кератиноцитами, макрофагами и эндотелиальными клетками при контактной реакции гиперчувствительности принадлежит цитокинам и простагландинам. Презентация антигена (1) вызывает каскадное выделение цитокинов (2). Это вначале приводит к активации и пролиферации Т-клеток CD4+(3), повышению экспрессии ICAM-1 и молекул МНС класса II на поверхности кератиноцитов и эндотелиальных клеток (4), а также к привлечению в кожу Т-клеток и макрофагов (3, 5). Последующая продукция ПГЕ кератиноцитами и макрофагами может ингибировать образование ИЛ-1 и ИЛ-2. В торможение реакции вносит вклад не только продукция ПГЕ, но и связывание активированных Т-клеток с кератиноцитами, а также ферментативное и клеточное разрушение комплекса гаптен-носитель.
Нанесение на кожу химических веществ может привести к усилению через 2 ч экспрессии ELAM-I и VCAM-1 и через 8ч — IСАМ-1, независимо от предварительной сенсибилизации. Большее значение по сравнению с ELAM-I и VCAM-I имеет IСАМ-1: эта молекула служит лигандом для LFA-1, присутствующего на поверхности лимфоидных и мислоидных клеток, и выполняет важную роль в удержании этих клеток в коже. Хемотаксические цитокины и «сигнальный эффект» перемещающихся клеток Лангерганеа привлекают Tx1-клетки. Т-клетки иммунологической памяти остаются в кожных капиллярах, где они могут инициировать реакцию и накапливаться независимо от присутствия специфического антигена.
Ряд цитокинов подавляет воспалительную реакцию
Через 48-72 ч реакция затухает; макрофаги и кератиноциты выделяют ПГЕ, который ингибирует продукцию ИЛ-1 и ИЛ-2; Т-клетки связываются с активированными кератиноцитами, и гаптеновый конъюгат подвергается ферментативной и клеточной деградации. Затуханию реакции способствуют следующие механизмы.
✵ Лимфокины, ингибирующие миграцию, препятствуют распространению воспалительной реакции.
✵ ТФРβ, выделяемый тучными клетками дермы, активированными кератиноцитами и лимфоцитами, ингибирует воспаление и блокирует пролиферативные эффекты ИЛ-1 и ИЛ-2.
✵ ИЛ-1, синтезируемый кератиноцитами после контакта с аллергенами, ингибирует окислительный метаболизм в макрофагах и тормозит продукцию ими провоспалительных медиаторов.
✵ ИЛ-10 ослабляет экспрессию молекул класса II и подавляет продукцию цитокинов и антиген-специфическую пролиферацию Тх1-клеток.
✵ Возможно участие внешних факторов; так, у мыши УФ-облучение индуцирует синтез специфического ингибитора активности ИЛ-1.
✵ Кератиноциты, экспрессирующие молекулы класса II, в отсутствие костимулирующих молекул не могут действовать как АПК, но, экспрессируя гаптен, способны вызывать клональную анергию Тх1-клеток.