ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 3. Лимфоидная система
ВТОРИЧНЫЕ ЛИМФОИДНЫЕ ОРГАНЫ И ОБРАЗОВАНИЯ
Из первичных лимфоидных органов образовавшиеся здесь лимфоциты перемещаются во вторичные, периферические лимфоидные ткани — плотно структурированные, инкапсулированные органы (селезенка и лимфатические узлы) и бескапсульные скопления. Не окруженные капсулой островки лимфоидной ткани, расположенные большей частью в слизистых оболочках, названы лимфоидной тканью слизистых оболочек (ЛТС).
Функции лимфоидной ткани слизистых оболочек отличаются от функций других органов лимфоидной системы в иммунном ответе
Селезенка отвечает на антигены, находящиеся в крови (в случае удаления этого органа у больного повышается восприимчивость к возбудителям, проникшим в кровоток). Лимфатические узлы защищают организм от антигенов, проникающих через кожу или слизистые оболочки и затем транспортируемых с лимфой по лимфатическим сосудам. Иммунный ответ на проникшие такими путями антигены складывается из секреции антител в кровоток и из местных клеточных реакций. В отличие от этого лимфоидная ткань слизистых оболочек защищает именно слизистые. В ЛТС происходит примирование, т. е. первый контакт иммунных клеток с антигеном, поступающим с поверхности эпителия. Лимфоидная ткань присутствует в слизистой оболочке кишечника, дыхательных путей (в частности, бронхов) и мочеполовых путей. Основной эффекторный механизм местного иммунного ответа на уровне слизистой оболочки — это секреция и транспорт секреторных антител класса IgA (sIgA) непосредственно на поверхность ее эпителия. Не удивительно, что большая часть лимфоидной ткани организма (>50%) находится в слизистых оболочках и особенно обильно представлена в кишечнике, поскольку через слизистые оболочки и проникают в основном антигены извне. По той же причине антитела IgA представлены в организме в наибольшем количестве относительно других изотипов антител.
Инкапсулированные вторичные лимфоидные органы
Селезенка. Селезенка расположена в левом верхнем квадранте брюшной полости, позади желудка и вплотную к диафрагме. У взрослого человека она имеет размеры примерно 13 х 8 см и массу 180—250 г. Снаружи селезенка покрыта соединительнотканной капсулой из пучков коллагеновых волокон, которые проникают в паренхиму органа, образуя короткие перекладины (трабекулы). Вместе с ретикулярной стромой они создают структурный каркас для массы заполняющих селезенку разнообразных клеток (рис. 3.5). В селезенке различают два основных типа ткани: красную пульпу и белую пульпу (мякоть).

Рис. 3.5. Гистологический срез селезенки, демонстрирующий ретикулярную строму красной пульпы. Окрашивание на ретикулин позволяет выявить кольцеобразно расположенные ретикулиновые волокна (К), которые поддерживают эндотелиальные клетки венозных синусоидов. Стенка этих сосудов образована несплошным слоем базальной мембраны - между эндотелиальными клетками имеются щели, которые обеспечивают свободное протекание плазмы в просвет сосуда, а также избирательный переход клеточных элементов из тяжей красной пульпы (КТ), х 125.
Белая пульпа. Белая пульпа состоит из лимфоидной ткани, образующей вокруг центральных артериол периартериолярные лимфоидные «муфты» (ПАЛМ) (рис. 3.6). В ПАЛМ имеются Т- и В-клеточные области: Т-клетки непосредственно окружают центральную артериолу, тогда как В-клетки могут образовывать первичные, «нестимулированные» фолликулы (агрегаты никогда не встречавшихся с антигеном лимфоцитов) или вторичные, «стимулированные» фолликулы, содержащие центры размножения с клетками иммунологической памяти (рис. 3.6 и 3.7). В центрах размножения присутствуют также фолликулярные дендритные клетки и фагоцитирующие макрофаги. В краевой зоне, расположенной над мантией, локализованы специализированные макрофаги и субпопуляции В-клеток, отвечающих на тимуснезависимые антигены II типа, например на полисахариды (см. гл. 11). Макрофаги и фолликулярные дендритные клетки в селезенке презентируют антигены В-клеткам. По входящим в краевую зону капиллярным ответвлениям центральной артериолы В-клетки и другие лимфоциты могут свободно поступать в ПАЛМ и покидать се. Отдельные субпопуляции лимфоцитов, в частности созревающие плазмобласты, могут проходить через краевую зону в красную пульпу по сосудистым перемычкам.
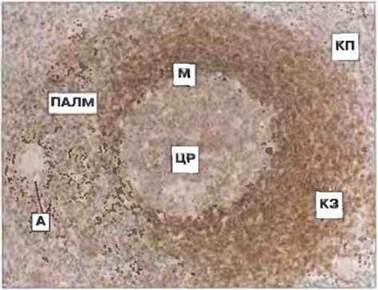
Рис. 3.6. Гистологический срез селезенки (лимфоидная ткань белой пульпы). Вторичный лимфоидный фолликул с центром размножения (ЦР) и мантией (М) окружен краевой зоной (КЗ) и красной пульпой (КП). Вблизи от фолликула проходит артериола (А), вокруг которой расположена периартериолярная лимфоидная муфта (ПАЛМ), состоящая в основном из Т-клеток. Краевая зона имеется лишь на одной стороне вторичного фолликула. (Фото любезно предоставлено проф. I. Maclennan.)
Красная пульпа. Эта ткань образована венозными синусоидами и клеточными тяжами (ретикулум), пространство между которыми заполняют оседлые макрофаги (рис. 3.8), эритроциты, тромбоциты, гранулоциты, лимфоциты и многочисленные плазматические клетки. Отметим, что помимо своих иммунологических функций селезенка выполняет функцию депонирования тромбоцитов, эритроцитов и гранулоцитов. В ней также разрушаются отжившие тромбоциты и эритроциты. Этот процесс, называемый гемокатерезом, протекает в красной пульпе. Осуществление депонирования и гемокатереза обеспечено особым строением кровеносной системы в селезенке (рис. 3.7). Окруженные ПАЛМ центральные артериолы оканчиваются капиллярами, которые свободно открываются в тяжах красной пульпы. Вследствие этого циркулирующие клетки, достигнув тяжей, задерживаются в них. Здесь макрофаги распознают и фагоцитируют отжившие тромбоциты и эритроциты. Не поглощенные и не разрушенные клетки крови возвращаются в кровоток, протискиваясь сквозь несплошную эндотелиальную выстилку венозных синусоидов (рис. 3.8) через щели между клетками, свободно пропускающие поток плазмы.
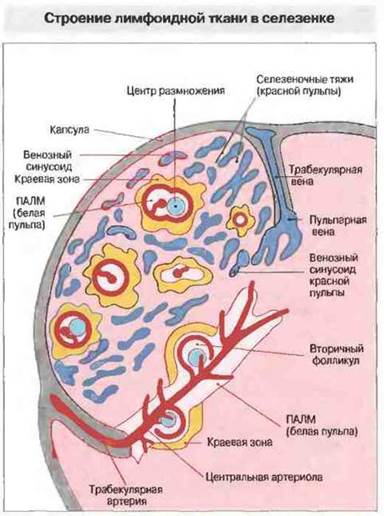
Рис. 3.7. Белая пульпа состоит из периартериолярных лимфоидных муфт (ПАЛМ), многие из которых содержат лимфоидные фолликулы. Она окружена краевой зоной, заполненной многочисленными макрофагами, антигенпрезентирующими клетками, медленно рециркулирующими В-лимфоцитами и нормальными киллерными клетками. В красной пульпе расположены венозные каналы (синусоиды), разделенные селезеночными тяжами. Кровь поступает в ткани селезенки по трабекулярным артериям, дающим начало ветвистым центральным артериолам. Часть этих артериол оканчивается в белой пульпе и питает центры размножения и краевую зону фолликула, но большинство достигает краевой зоны или соседних с нею участков. Некоторые ветви артериол заходят непосредственно в красную пульпу, оканчиваясь в селезеночных тяжах. Из венозных синусоидов кровь собирается в пульпарные вены, затем в трабекулярные вены и из них в селезеночную вену.
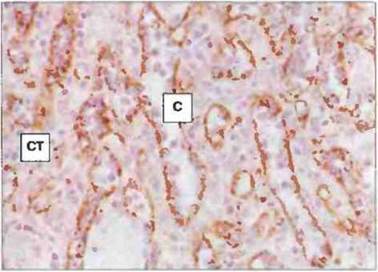
Рис. 3.8. Венозные синусоиды и тяжи красной пульпы. Иммуноферментное окрашивание катепсина D выявляет макрофаги в стенках синусоида (С) и в селезеночных тяжах (СТ). (Фото любезно предоставлено д-ром А. Stevens и проф. J. Lowe.)
Лимфоузлы и лимфатическая система. Лимфатические узлы составляют часть системы, которая «вылавливает» антигены из тканевой жидкости и лимфы во время ее протекания от периферии к грудному протоку через главные лимфатические коллекторы (рис. 3.9). Лимфоузлы обычно расположены в местах разветвления лимфатических сосудов. В стратегических пунктах системы — шейной, подмышечной и паховой областях, средостении и брюшной полости — они образуют скопления, собирающие лимфу из соответствующих поверхностных и глубоких областей тела. Лимфоузлы, расположенные поверхностно и называемые подкожными, защищают кожу. Висцеральные (глубокие) лимфоузлы осуществляют защиту слизистых оболочек дыхательных путей, пищеварительного тракта и мочеполовых путей.
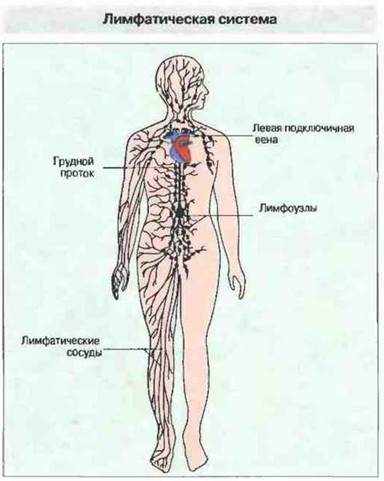
Рис. 3.9. Лимфоузлы находятся в местах слияния лимфатических сосудов и образуют сеть, которая собирает и фильтрует интерстициальную тканевую жидкость. Они подразделяются на подкожные и висцеральные, т. е. дренирующие глубокорасположенные ткани и внутренние органы тела. В конечном итоге лимфа достигает грудного лимфатического протока, впадающего в левую подключичную вену, и таким образом возвращается в кровоток.
Лимфатические узлы человека — это образования округлой или бобовидной формы, диаметром 2—8 мм, с углублением для входа и выхода кровеносных сосудов, называемым воротами. Лимфа поступает в узел по нескольким приносящим (афферентным) лимфатическим сосудам и выходит из него по единственному выносящему (эфферентному) лимфатическому сосуду через ворота. Снаружи лимфоузел покрыт капсулой из коллагеновых волокон (рис. 3.10). Радиально расположенные перегородки — трабекулы - вместе с тяжами ретикулярного остова поддерживают заполняющие узел разнообразные клетки. В лимфоузле различают В-клеточную корковую область, или кортекс, Т-клеточную (паракортикальную) область и центральную (мозговую) область (рис. 3.11 и 3.12). Последняя образована клеточными тяжами, содержащими Т- и В-лимфоциты, плазматические клетки и макрофаги.
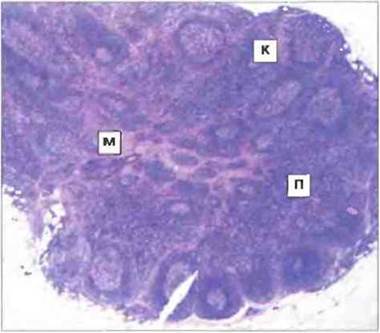
Рис. 3.10. Гистологический срез лимфоузла. Лимфоузел окружен соединительнотканной капсулой и имеет три основные области: В-клеточную корковую, или кортекс (К), Т-клеточную паракортикальную (П) и мозговую (М), образованную тяжами лимфоидной ткани (Т- и В-клеточная область, богатая плазматическими клетками и макрофагами). Окрашивание гематоксилином/эозином. х 10. По Zucker-Franklin D., Greaves V.F., Grossi С.Е. et al. 1988. Atlas of Blood Cells: Function and Pathology. Vol. II. 2nd edn. Philadelphia: Lea and Febiger.
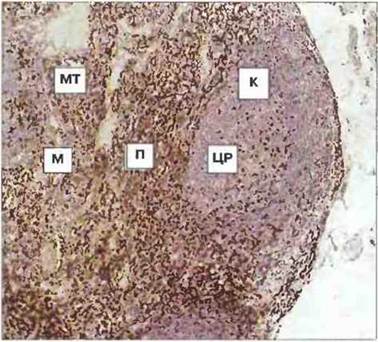
Рис. 3.11. Гистологическая структура лимфоузла. Видны корковая (К), паракортикальная (П) и мозговая (М) области. Срез окрашен для выявления локализации Т-клеток. Больше всего их в паракортикальной области и некоторое количество присутствует в центре размножения (ЦР) вторичного лимфоидного фолликула, в корковой области и мозговых тяжах (МТ). (Фото любезно предоставлено д-ром A. Stevens и проф. J. Lowe.)
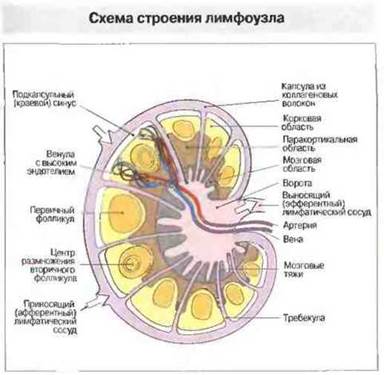
Рис. 3.12. Под капсулой, образованной коллагеновыми волокнами, находится подкапсульный синус, выстланный эндотелием и макрофагами. По приносящим лимфатическим сосудам из межклеточного пространства окружающих тканей и смежных лимфоузлов в него поступают лимфоциты и антигены. Корковая область заполнена в основном В-клетками, образующими первичные, а чаще всего вторичные (т. е. содержащие центры размножения) фолликулы. Паракортикальная область содержит главным образом Т-клетки. Каждый лимфоузел снабжен собственными артерией и веной Поступление лимфоцитов из кровотока происходит в паракортикальной области по функционально специализированным венулам с высоким эндотелием (ВЭВ). В мозговой области содержится не только большая часть плазматических клеток лимфоузла, но также Т- и В -клетки, образующие тяжи лимфоидной ткани. Лимфоциты покидают лимфоузел через выносящий лимфатический сосуд.
Паракортикальная область содержит много переплетенных отростками (интердигитатных) клеток, экспрессирующих в большом количестве поверхностные антигены МНС класса II. Эти клетки собираются здесь, мигрируя из кожи (клетки Лангерганса) или из слизистых оболочек (дендритные клетки) и транспортируя при этом в лимфоузлы процессированные антигены из наружных покровов тела и слизистых оболочек (рис. 3.13). Основная масса лимфоидной ткани лимфатического узла сосредоточена в корковой и пара кортикальной областях. Мозговая область образована тяжами, которые разграничивают лимфатические (мозговые) синусы, собирающие лимфу в краевой синус и далее в выносящий лимфатический сосуд (рис. 3.12). Вдоль синусов, большей частью в мозговой области, расположены клетки, фагоцитирующие детрит. В процессе протекания лимфы через лимфоузел из приносящих лимфатических сосудов в выносящий, эти фагоцитарные клетки вылавливают из нее корпускулярные антигены и транспортируют их в собственно лимфоидную ткань лимфатического узла.
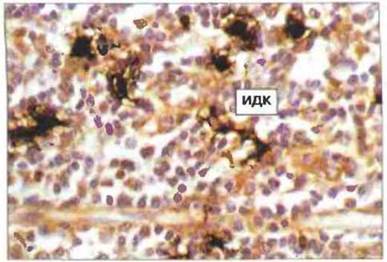
Рис. 3.13. Разветвленные, переплетенные отростками (дендритные интердигитатные) клетки (ИДК) в паракортикальной области лимфоузла контактируют между собой и с Т-лимфоцитами. (Фото любезно предоставлено д-ром A. Stevens и проф. J. Lowe.)
В корковой области содержатся скопления В-клеток, образующих первичные и вторичные фолликулы, тогда как в пара кортикальной области находятся главным образом Т-клетки. Вследствие этого после инъекции любого Т-зависимого антигена в кожу или слизистую оболочку (см. м. II) в паракортикальной области лимфоузла, дренирующего место введения, наблюдается активная пролиферация Т-клеток. Еще одно доказательство именно такой локализации Т клеток демонстрируют больные с наследственной аплазией тимуса (синдром Ди Джорджи), у которых паракортикальные области лимфоузлов содержат меньше клеток, чем в норме. Подобные явления наблюдаются у неонатально тимэктомированных или наследственно бестимусных (голых) мышей и крыс.
Центры размножения можно обнаружить во вторичных фолликулах лимфоузлов, стимулированных антигеном. Они сходны с центрами размножения в В-клеточных областях селезеночных ПАЛМ и ЛТС. Большие и малые клетки фолликулярных центров размножения названы центробластами и центроцитами. У пролиферирующих В клеток в центрах размножении ядро имеет характерную расщепленную форму, что служит ценным признаком для дифференциальной диагностики некоторых злокачественных лимфопролиферативных заболеваний, таких как центробластно-цеитроцитарные лимфомы, возникающие из этих клеток.
Центры размножения окружены мантией из лимфоцитов (рис. 3.14). На В-клетках в зоне мантии сопряженно экспрессируются (коэкспрессия) поверхностные IgM и IgD (рис. 3.15). Мантия большинства вторичных фолликулов имеет утолщение (корону) со стороны капсулы лимфоузла (рис. 3.15). Кроме В-клеток, во вторичных фолликулах содержатся фолликулярные деидритные клетки (ФДК) (рис. 3.16), некоторое количество макрофагов (рис. 3.17) и небольшое число Т-клеток CD4+, которые взаимодействуют с дендритными клетками центров размножения (см. гл. 2). Все перечисленные клетки вместе со специализированными макрофагами краевого синуса способствуют, по-видимому, возникновению В-клеточного иммунного ответа, в частности развитию иммунологической В-клеточной памяти — одной из главных функций центров размножения (более подробно клеточное строение центра размножения изложено в гл. 12).
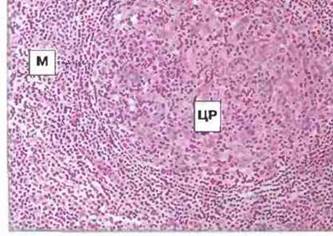
Рис. 3.14. Структура вторичного фолликула лимфатического узла. Крупный центр размножения (ЦР) окружен мантией (М).
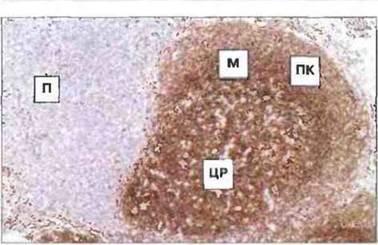
Рис. 3.15. Распределение В-клеток в корковой области лимфоузла. Иммуногистохимическое окрашивание поверхностных иммуноглобулинов выявило В-клетки, в наибольшем числе сконцентрированные во вторичном фолликуле - центре размножения (ЦР) и мантии (М), а также между фолликулом и капсулой узла, в подкапсульной зоне (ПК). Некоторое количество В-клеток присутствует в паракортикальной области (П), состоящей в основном из Т-клеток (см. рис. 3.11).

Рис. 3.16. Срез вторичного лимфоидного фолликула, демонстрирующий сеть из дендритных клеток (окрашивание с помощью конъюгированных с ферментом моноклональных антител, специфичных к ФДК).
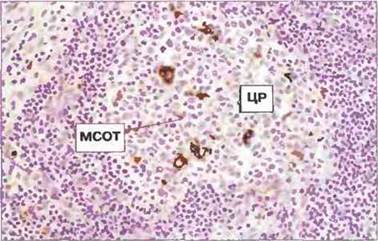
Рис. 3.17. Макрофаги центра размножения. Иммуноферментное окрашивание, специфичное для катепсина D, выявило несколько макрофагов, локализованных в центре размножения (ЦР) вторичного фолликула. В случае, когда эти клетки фагоцитируют апоптотичные В-лимфоциты, их называют макрофагами, содержащими окрашенные тельца (остатки В-клеточных ядер) (МСОТ).
Лимфоидная система слизистых оболочек
Лимфоидная ткань слизистых оболочек. Скопления бескапсульной лимфоидной ткани можно обнаружить в собственной пластинке слизистых оболочек и в подслизистой ткани желудочно-кишечного тракта, дыхательных и мочеполовых путей (рис. 3.1). Лимфоидные клетки образуют в них одиночные или агрегированные скопления с центрами размножения (вторичные фолликулы). Так, в миндалинах человека, где содержание лимфоидной ткани значительное, часто присутствуют крупные вторичные фолликулы; они перемежаются Т-клеточными зонами, через которые проходят венулы с высоким эндотелием. Миндалины, названные в соответствии с их расположением нёбными, глоточными (аденоиды) и язычными, образуют кольцо Вальдейера (рис. 3.1). Гистологический срез язычной миндалины представлен на рис. 3.18. Аналогичные лимфоидные образования прилежат к слизистой оболочке, выстилающей бронхи и мочеполовые пути. Сами слизистые оболочки пищеварительной, дыхательной и мочеполовой систем содержат дендритные клетки, необходимые для поглощения, процессинга и транспорта антигенов в регионарные лимфоузлы. Скопления лимфоидной ткани, расположенные в собственной пластинке слизистой оболочки желудочно-кишечного тракта, часто распространяются в подслизистый слой. Они имеют форму одиночных фолликулов (рис. 3.19) или сгруппированных узелков, как, например, в червеобразном отростке слепой кишки (рис. 3.20). Пейеровы бляшки обычно встречаются в нижней части подвздошной кишки. Покрывающий их эпителий кишечника (эпителий, ассоциированный с фолликулами) способен транспортировать антигены и микробы в лимфоидную ткань. Эту специализированную функцию выполняют особые эпителиальные клетки, рассеянные среди энтероцитов; они названы М-клетками, поскольку их обращенная в просвет кишечника поверхность образует многочисленные микроскладки (рис. 3.21). В базолатеральной области М-клеток имеются глубокие инвагинации плазматической мембраны — карманы, в которых располагаются В- и Т-лимфоциты, дендритные клетки и макрофаги (рис. 3.22). Антигены и микробы подвергаются трансцитозу в эти карманы и далее в организованную субэпителиальную лимфоидную ткань слизистой оболочки. М-клетки встречаются не только в участках нейеровых бляшек, но и в других лимфоидных образованиях слизистых оболочек.
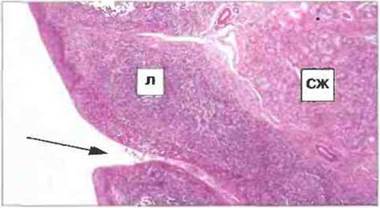
Рис. 3.18. Структура язычной миндалины. Язычная миндалина расположена в задней трети языка и состоит из скоплений лимфоидной ткани (Л), которая содержит крупные вторичные фолликулы и ассоциирована со слизистой оболочкой, образующей в этом участке глубокие щелеподобные инвагинации (стрелка). Вокруг миндалины видны содержащие слизь слюнные железы (СЖ). Такую же структуру имеют все прочие миндалины. (Фото любезно предоставлено д-ром A. Stevens и проф. J. Lowe.)
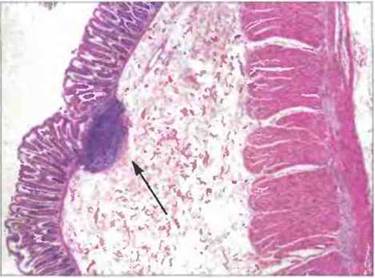
Рис. 3.19. Одиночный лимфоидный фолликул в толстой кишке. Этот узелок лимфоидной ткани расположен в слизистой оболочке и в подслизистой основе кишечной стенки (стрелка) (Фото любезно предоставлено д-ром A. Stevens и проф. J. Lowe.)
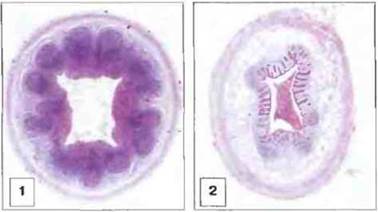
Рис. 3.20. Лимфоидные узелки в червеобразном отростке у человека. 1. Аппендикс десятилетнего ребенка; крупные лимфоидные узелки в слизистой оболочке, распространяющиеся в подслизистую основу. 2. Аппендикс мужчины 36 лет; заметно существенное сокращение лимфоидной ткани с фактически полным исчезновением фолликулов - возрастная атрофия лимфоидной ткани, характерная не только для червеобразного отростка. (Фото любезно предоставлено д-ром A. Stevens и проф. J. Lowe.)
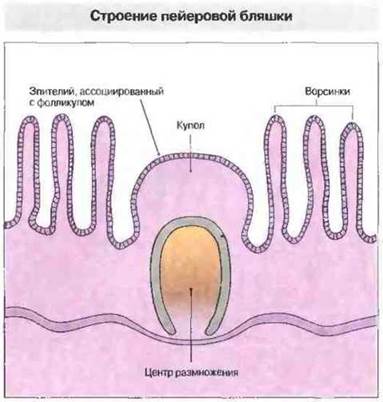
Рис. 3.21. Куполообразный выступ, образуемый слизистой оболочкой кишечника, на участке, лишенном ворсинок. Поверхностный эпителий в этом участке, называемый эпителием, ассоциированным с фолликулами (ЭАФ), содержит М-клетки. В глубине слизистой оболочки расположено скопление вторичных лимфоидных фолликулов с крупными центрами размножения. Окружающие тимус-зависимые межфолликулярные зоны содержат интердигитатные клетки и венулы с высоким эндотелием. Область купола между ЭАФ и фолликулами заполнена преимущественно В-клетками, большинство которых относится к клеткам иммунологической памяти.
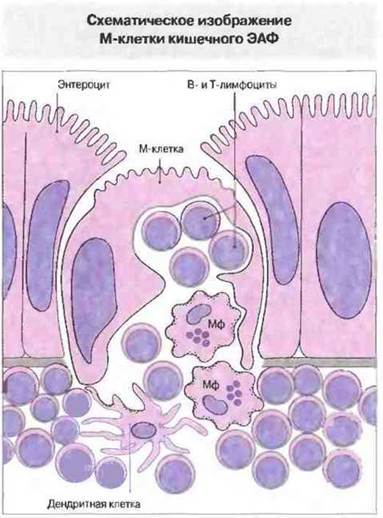
Рис. 3.22. Во внутриклеточном кармане находятся лимфоциты и нередко макрофаги (Мф). Эндоцитированные М-клеткой антигены попадают через этот карман в субэпителиальную лимфоидную ткань. (ЭАФ - эпителий, ассоциированный с фолликулами.)
При местном гуморальном иммунном ответе на уровне слизистой оболочки происходит образование антител в основном изотипа IgA. Секреторные IgA — это антитела, способные проникать через мембраны эпителиальных клеток для обеспечения зашиты от патогенных микробов (рис. 3.23).
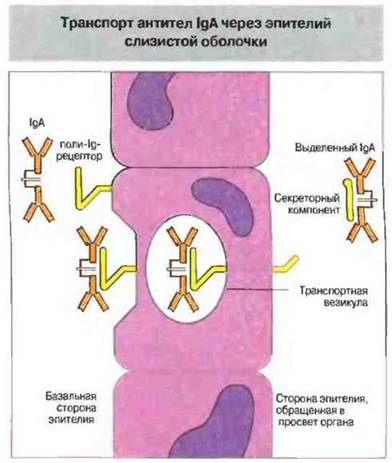
Рис. 3.23. Димеры IgA (sIgA), выделяемые плазматическими клетками в собственной пластинке слизистой оболочки кишечника, связываются поли-Ig-рецепторами на базальной стороне эпителия. Затем комплексы sIgA-рецептор путем эндоцитоза и переноса через клетку в транспортных везикулах (с мембраной которых они связаны) доставляются на поверхность эпителия, обращенную в просвет кишечника. Здесь транспортные везикулы сливаются с плазматической мембраной клеток эпителия, высвобождая димеры IgA с присоединенным секреторным компонентом (фрагмент рецептора) в просвет, где секреторный компонент предохраняет димеры IgA от расщепления протеолитическими ферментами.
Лимфоциты слизистых оболочек. Дополнительно к организованной лимфоидной ткани, образующей единую систему ЛТС, в слизистой оболочке желудка, кишечника, верхних и нижних дыхательных путей и некоторых других органов присутствует множество рассеянных лимфоцитов и плазматических клеток. Лимфоциты обнаруживаются в соединительной ткани собственной пластинки и в эпителиальной выстилке.
Среди лимфоцитов собственной пластинки (ЛСП) преобладают активированные Т-клетки, но в значительном количестве присутствуют также активированные В-клетки и плазмоциты. Эти плазматические клетки секретируют антитела преимущественно изотипа IgA, транспортируемые через клетки эпителия и высвобождаемые в просвет органа (рис. 3.23).
Внутриэпителиальные лимфоциты (ВЭЛ) представлены в основном Т-клетками, отличными по фенотипу от ЛСП (рис. 3.24).
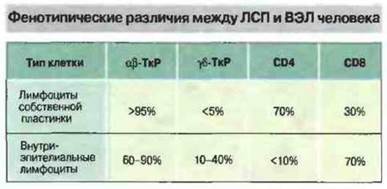
Рис. 3.24. Фенотипические маркеры лимфоцитов собственной пластинки слизистой оболочки кишечника (ЛСП) в основном сходны с маркерами лимфоцитов периферической крови. Значительная часть внутриэпителиальных лимфоцитов (ВЭЛ) относится к γδ-Т-клеткам, многие из которых экспрессируют CD8, вероятно как маркер активации.
Большинство ЛСП и ВЭЛ относятся к клеткам иммунологической памяти, несущим маркер CD45RO. Они слабо отвечают па стимуляцию антителами к CD3, но чувствительны к другим механизмам активации, например опосредованным CD2 или CD28.
Покоящиеся Т-клетки периферической крови не экспрессируют интегриновую α-цепь HML-1 (СD103), но стимуляция фитогемагглютинином (ФГА) индуцирует ее синтез. Антитела к этой цепи митогенны для тех же клеток и вызывают у них экспрессию α-цепи низкоаффинных рецепторов ИЛ-2 (CD25). Полипептид HML-1 это α-цепь из семейства интегринов, образующая при взаимодействии с β7-иепью гетеродимер αHML-1-β7— интегрин, который синтезируют ВЭЛ и другие активированные лимфоциты.
Внутриэпителиальные лимфоциты выделяют ряд цитокинов, в том числе ИФγ и ИЛ-5. Одна из функций ВЭЛ состоит, предположительно, в иммунологическом надзоре, направленном на устранение мутантных иди инфицированных вирусами клеток.