ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 3. Лимфоидная система
ЦИРКУЛЯЦИЯ ЛИМФОЦИТОВ
Миграция лимфоцитов из первичных во вторичные лимфоидные ткани уже описана выше. Оказавшись во вторичных лимфоидных органах и образованиях, многие лимфоциты не остаются в них, а перемещаются из одного лимфоидного органа в другой по кровеносным и лимфатическим сосудам (рис. 3.25).
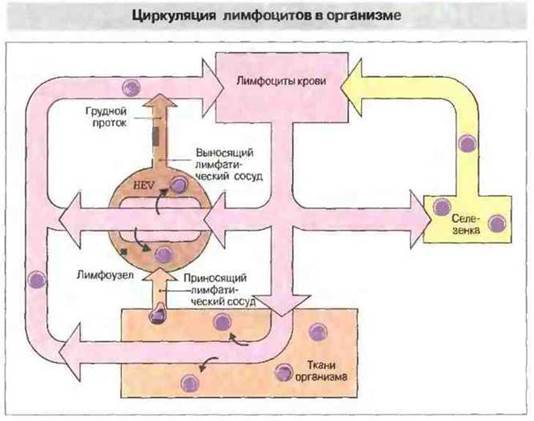
Рис. 3.25. Лимфоциты из кровотока проникают в лимфоузлы и в ЛТС через специализированный высокий эндотелий посткапиллярных венул (ВЭВ); затем они покидают лимфоидную ткань по выносящим лимфатическим сосудам и, пройдя сквозь другие лимфоузлы, возвращаются в кровоток по грудному протоку, впадающему у человека в левую подключичную вену. В селезенке лимфоциты входят в белую пульпу через краевые зоны; затем, попав в синусоиды красной пульпы, покидают орган через селезеночную вену.
Лимфоциты выходят из кровотока в лимфоидную ткань через стенки венул с высоким эндотелием. Некоторые лимфоциты переходят из кровотока в лимфоидную ткань через обычные посткапиллярные венулы, однако у большинства млекопитающих этот переход осуществляется преимущественно через специализированные участки венозного русла - венулы с высоким эндотелием, или высокоэндотелиальные венулы (ВЭВ) (рис. 3.26 и 3.27). В лимфоузлах эти сосуды находятся главным образом в паракортикальной области и иногда в корковой, но не в мозговой. Вместе с тем часть лимфоцитов, в первую очередь Т-клетки, поступают в регионарный лимфоузел из дренируемой им области но приносящим лимфатическим сосудам, а не по ВЭВ. Тем же путем в лимфоузлы поступает большинство антигенов. Венулы с высоким эндотелием обнаружены не только в лимфоузлах, но и в ЛТС, а также в тимусе.
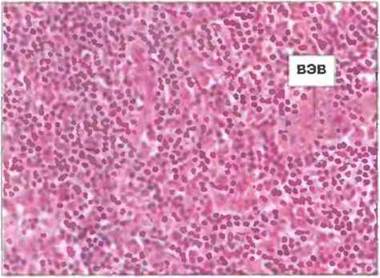
Рис. 3.26. Венула с высоким эндотелием (ВЭВ) в паракортикальной области лимфоузла. Лимфоциты выходят из кровотока через высокий эндотелий (ВЭВ) и проникают в лимфоузел. Окрашивание гематоксилином/эозином. х 200. (Фото любезно предоставлено д-ром A. Stevens и проф. J. Lowe.)
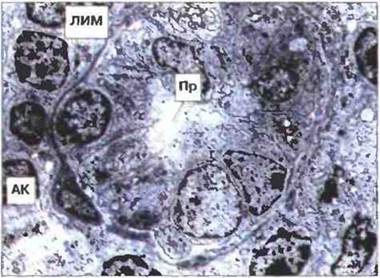
Рис. 3.27. Электронная микрофотография венулы с высоким эндотелием в паракортикальной области лимфоузла. Вблизи базальной мембраны виден лимфоцит (ЛИМ), покидающий просвет (Пр) венулы, которая снаружи частично охвачена адвентициальной клеткой (АК). х 1600.
Венулы с высоким эндотелием управляют циркуляцией лимфоцитов. Эти венулы выстланы кубическими эндотелиальными клетками, которые в отличие от покоящихся плоских клеток эндотелиальной выстилки обычных венул экспрессируют при активации разнообразные молекулы межклеточной адгезии. Один из механизмов активации клеток эндотелия опосредован локально синтезируемыми цитокинами, такими как ИФγ, ИЛ-1 и ФИО.
Эндотелий обычных венул может превращаться в кубический в участках хронического воспаления, например в коже или в синовиальных оболочках, где ВЭВ в норме отсутствуют. Появившиеся ВЭВ направляют в очаг воспаления специализированные субпопуляции Т-лимфоцитов. Активированные кубические эндотелиоциты экспрессируют ряд молекул межклеточной адгезии из суперсемейства иммуноглобулинов [1CAM-1 (CD54), 1САМ-2 (CD102) и VCAM-1 (CD106)] или из семейства селектинов, в том числе Е-селектин [ELAM-1 (CD62E)] и Р-селектин (CD62P). Р-селектин хранится втельцах Вейбеля-Палала эндотелиальных клеток кровеносных капилляров и при активации быстро доставляется на поверхность эндотелиоцита (см. гл. 5). Ведущая роль в прилипании лимфоцитов к эндотелию принадлежит CD44 — белку с мол. массой 90 кДа, который экспрессируют все лейкоциты. Предположительно, между лимфоцитами и эндотелием возникают специфические лиганд-рецепторные взаимодействия, в результате которых лимфоциты направляются в определенные ткани-мишени. Это происходит за счет экспрессии эндотелием специфичных для данного органа «адресатов», например MAdCAM-1 на эндотелиоцитах в кишечнике и VCAM-I на эндотелиальных клетках в других органах. Для избирательного органоспецифического распределения лимфоцитов важны также особые молекулы «хоминга» (от англ, homing — возврат). Например, решающее значение для возвращения лимфоцитов в лимфоидную ткань кишечника имеют их интегрины α4β7, которые связываются с алрессинами MAdCAM-1 на эндотелиальных клетках ВЭВ в пейеровых бляшках. У мышей, дефектных по гену β7, образование лимфоидной ткани кишечника существенно нарушено.
Благодаря рециркуляции любой антиген экспонируется множеству лимфоцитов. Из лимфоузлов лимфоциты возвращаются в кровоток по выносящим лимфатическим сосудам, через грудной проток и подключичную вену. Ежечасно в рециркуляцию вовлекается 1—2% лимфоцитов. В итоге этот процесс позволяет множеству антигенспецифичных лимфоцитов встретиться с соответствующими антигенами, проникшими в их микроокружение в периферических лимфоидных органах. Особая важность рециркуляции становится очевидной, если вспомнить, что лимфоидные клетки моноспецифичны, и лишь ограниченное число лимфоцитов способно распознавать каждый конкретный антиген.
В норме рециркуляция лимфоцитов через лимфоузлы происходит постоянно, но если в лимфоидную ткань ранее сенсибилизированного к тому или иному антигену животного повторно попадает данный антиген, рециркуляция прекращается приблизительно на 24 ч. Временная остановка рециркуляции обусловлена в этом случае избирательной задержкой антига (специфичных лимфоцитов в лимфоузлах, дренирующих место проникновения антигена. Например, образовавшиеся в результате контакта с антигеном лимфобласты уже не рециркулируют, оставаясь, по-видимому, в участке встречи с антигеном.
Лимфоидная ткань, ассоциированная со слизистыми оболочками, отличается как система от других лимфоидных органов в том числе и тем, что лимфоциты ЛТС возвращаются в процессе рециркуляции главным образом в эту ткань. Так, лимфоциты, стимулированные в пейеровых бляшках, проходят через регионарные лимфоузлы в кровоток, а затем возвращаются «домой», в собственную пластинку слизистой оболочки кишечника (рис. 3.28). Такая специфическая рециркуляция объясняется тем, что эти лимфоциты эспрессируют молекулы «возврата домой», которые связываются со специфическими молекулами адгезии - адресатами — на поверхности эндотелиоиитов. Адресаты экспрессирует только эндотелий венул лимфоидной ткани слизистых оболочек, но не ВЭВ обычных лимфоузлов (см. выше), что и обеспечивает избирательную рециркуляцию. По той же причине стимуляция антигеном в области слизистой оболочки (в том или ином участке организма) вызывает системное образование антител преимущественно в ЛТС.
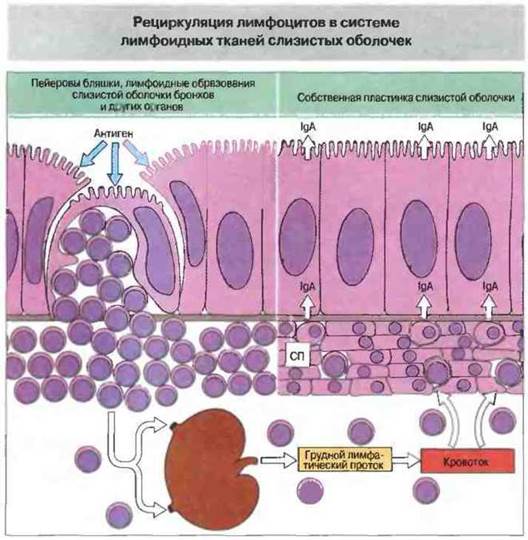
Рис. 3.28. Лимфоидные клетки, активированные антигеном в пейеровых бляшках (или в лимфоидных образованиях, ассоциированных с бронхами и другими слизистыми оболочками) через регионарные лимфоузлы и затем по грудному протоку мигрируют в кровяное русло, из которого вновь возвращаются в собственную пластинку (СП) эпителия кишечника и других слизистых оболочек, вблизи места антигенного примирования или на значительном удалении от него. Таким образом лимфоциты, стимулированные на слизистой оболочке в одной из частей тела, могут избирательно распределяться по системе ЛТС. Избирательность расселения достигается благодаря экспрессии особых молекул межклеточной адгезии на лимфоцитах и на клетках ВЭВ слизистых оболочек.
Вопросы для размышления
■ Почему млекопитающие нуждаются в столь высокоспециализированной лимфоидной системе, тогда как более примитивные животные способны выживать без нее?
■ Насколько важна рециркуляция лимфоцитов в системе лимфоидной ткани слизистых оболочек и какие молекулы обеспечивают ее избирательность?
■ В чем сходство и различия между лимфоидной тканью слизистых оболочек и остальными органами лимфоидной системы? Важна ли для выживания одна из этих частей лимфоидной системы более, чем другая?
■ Чем лимфоидная система отличается от любой другой системы организма, например от дыхательной или репродуктивной? (Возможно, для ответа вам понадобится освежить знания о различных системах организма по учебнику физиологии.)
■ Какие вспомогательные клетки присутствуют в лимфоидных тканях и в чем их функции? В чем состоят функции первичных и вторичных лимфоидных органов и образований?
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Butcher Е.С., Picker L.J. 1996. Lymphocyte homing and homeostasis. Science 272: 6—66.
Hogg N.. Berlin C. 1995. Structure and function of adhesion receptors in leukocyte trafficking. Immunol. Today 16. 327-330.
Kuby J. 1997. Immunology. 3rd edn. Cells and Organs of the Immune System. New York: W.H. Freeman and Co. 47-83.
Playfair J.H.L. Immunology at a Glance. 6th edn.
Oxford: Blackwell Scientific Publications. 1996.
Roitt I.M. Essential Immunology. 9th edn. Oxford: Blackwell Scientific Publications, 1997.