ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 13. Регуляция иммунного ответа
РОЛЬ ЛИМФОЦИТОВ В ИММУНОРЕГУЛЯЦИИ
Т-лимфоциты оказывают очевидное положительное влияние на иммунный ответ, выполняя хелперную функцию. Кроме того, в зависимости от типа хелперных Т-клеток (Tx1 или Тх2) иммунный ответ может носить характер гуморального или клеточного. Имеется также убедительное доказательство того, что Т-клетки способны подавлять иммунный ответ (рис. 13.8).
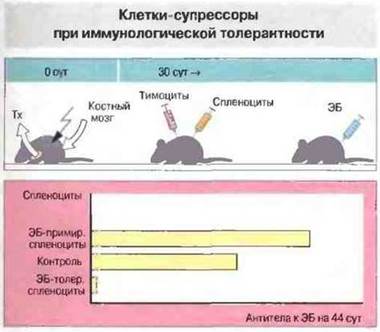
Рис. 13.8. Тимэктомированным и облученным мышам вводили клетки костного мозга, после чего через 30 сут - тимоциты и спленоциты с одновременной иммунизацией эритроцитами барана (ЭБ). На 44 сутки у реципиентов, получивших спленоциты животных, примированных иммуногенной дозой ЭБ (ЭБ-примированные спленоциты), зарегистрирован эффективный иммунный ответ (антителообразование), У животных, которым спленоциты не вводили (контроль), реакция была умеренной. В варианте с введением клеток толерантных животных (ЭБ-толерантные спленоциты; толерантность индуцировали высокой дозой ЭБ) антителообразование отсутствовало. Это показывает, что клетки толерантных животных активно подавляли иммунный ответ у реципиентов.
Т-клетки CD4+могут предотвращать возникновение аутоиммунитета
При многих экспериментальных аутоиммунных заболеваниях Т-клетки CD4+, образующиеся после введения высоких доз аутоантигенов (часто в растворимой или дезагрегированной форме), предотвращают последующую индукцию аутоиммунного процесса. Так, Т-клетки CD4+предотвращают образование аутоантител к тиреоглобулину (рис. 13.9).
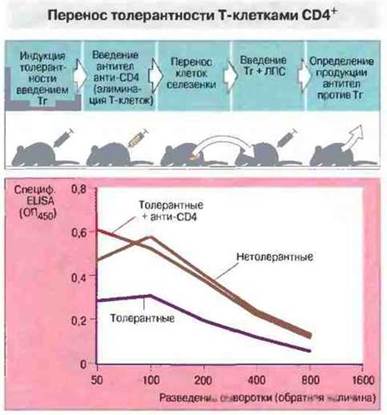
Рис. 13.9. Мышам вводили 20 мкг мышиного тиреоглобулина (Тг) для индукции толерантности (контроль - без индукции толерантности). Половине толерантных животных затем инъецировали in vivoэлиминирующие антитела анти-СБ4 для истощения пула Т-клеток CD4+. Клетки селезенки каждой мыши из этих трех групп (нетолерантные мыши; толерантные мыши; толерантные мыши, получившие анти-СD4) переносили облученному сингенному реципиенту. Затем реципиентам вводили мышиный Тг и ЛПС, после чего определяли продукцию антител против Тг при помощи иммуноферментного анализа (ELISA, см. гл. 29). Обработка антителами анти-СD4 устраняла перенос толерантности.
Установлено также, что одновременное введение мышам антител анти-СD4 (блокирующих презентацию антигена, опосредованную МГ1С-молекулами класса II) и иммуногенной дозы тиреоглобулпна не только предотвращает развитие аутоиммунитета, но приводит к образованию популяции Т-клеток CD4+, способных передать специфическую толерантность непримированным мышам-реципиентам (рис. 13.10). Механизм, посредством которого Т-клетки осуществляют такого рода отрицательное влияние, не вполне ясен. Однако проведенные недавно эксперименты указывают на то, что частично или полностью подавлять иммунный ответ могут продуцируемые Тх-клетками цитокины ТФРβ, ИЛ-4 и ИЛ-10.
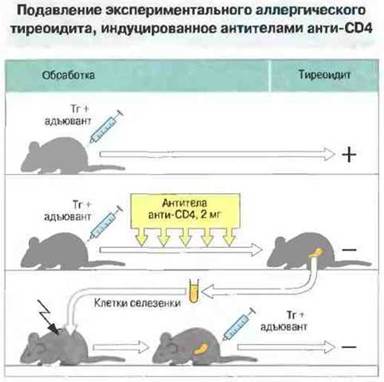
Рис. 13.10. У мышей, иммунизированных 50 мкг мышиного тиреоглобулина (Тг), возникал тиреоидит и появлялись антитела анти-Тг. Если на протяжении 11 сут после иммунизации мышам вводили неэлиминирующие моноклональные антитела анти-СD4 для блокирования взаимодействия CD4 с молекулами МНС класса II, тиреоидит не развивался. Перенос клеток селезенки от этих мышей облученным животным предотвращал возникновение у них тиреоидита после иммунизации Тг. (У иммунизированных мышей, получивших контрольные Т-клетки, заболевание возникало.)
В регуляции антителообразования участвуют различные субпопуляции Тх-клеток
Некоторые наблюдения, касающиеся регуляции синтеза IgE-антител, можно объяснить тем, что Тх-клетки CD4+ разных субпопуляций продуцируют разные цитокины. Между отдельными субпопуляциями Тх-клеток существует взаимосвязь в виде перекрестной регуляции их активности; секретируемый Tx1-клетками ИФγ способен ингибировать реактивность Тх2-лимфоиитов; ИЛ-10, образуемый Тх2-клетками, понижает экспрессию молекул В7 и ИЛ-12 антигенпрезентирующими клетками, что в свою очередь ингибирует активацию Тх1 -лимфоцитов. Кроме того, на экспрессию высокоаффинного рецептора ИЛ-12 (ИЛ-12Р), необходимого для развития Тх1-клеток, влияет баланс цитокинов. Высокоаффинный ИЛ-12Р содержит в своей структуре две цепи, β1и β2, причем обе они вместе экспрессируются только клетками Tx1. Цепь β1 имеют и Тх1-, и Тх2-лимфоциты, а экспрессию β2-цепи индуцирует ИФγ и ингибирует ИЛ-4. На развитие субпопуляций Т-клеток оказывает также влияние ИФα, который благоприятствует формированию субпопуляций Tx1-лимфоцитов даже в присутствии ИЛ-4 и в условиях нейтрализации ИЛ-12. Таким образом, преимущественная активация Tx1- или Тх2-клеток может приводить к иммунному отклонению - избирательному развитию эффекторного ответа определенного типа. Такой избирательный сдвиг ответа может быть использован в терапии аутоиммунных и аллергических заболеваний.
Адоптивным переносом Т-клеток CD8+ можно вызвать резистентность и толерантность
Установлено, что Т-клетки CD8+также регулируют иммунный ответ. В селезенке животных, у которых индуцировали толерантность к ОБМ путем перорального введения антигена (см. выше), были обнаружены Т-лимфоциты CD84, способные при адоптивном переносе создавать у реципиентов резистентность к ЭАЭ. Эти Т-клетки не только супрессируют Т-клеточный ответ на ОБМ invitro, но могут также вызывать эффект «подавления свидетеля» в отношении других, неродственных антигенов. Предположительно данный эффект опосредован ТФРβ.
Регуляция иммунного ответа Тх2-клетками CD4+представляет собой нормальный физиологический процесс
Роль регуляторных эффектов, опосредованных Т-клетками CD4+или CD8+, в обычных физиологических условиях остается под вопросом. Однако обнаружение в норме у животных Т-лимфоцитов CD4+, способных предупреждать развитие аутоиммунитета, свидетельствует об их значении в поддержании нормального гомеостаза. Кроме того, для крысы и мыши установлено, что после удаления Тх2-клеток СD4+, продуцирующих в обычных условиях ИЛ-4 и ИЛ-10, нарушается регуляция иммунного ответа. Этот факт убедительно доказывает, что регуляция иммунного ответа, осуществляемая Тх2-лимфоцитами CD4 (но не Tx1-клетками CD4+), представляет собой нормальный физиологический процесс, а не артефакт (рис. 13.11). По имеющимся данным, при возникновении колита у мышей (рис. 13.11) регулирующую роль в развитии патологического процесса может играть ТФРβ. Это согласуется с результатами опытов по изучению колита у мышей, лишенных гена ТФРβ методом генного нокаута.

Рис. 13.11. Рисунок иллюстрирует частоту возникновения колита у мышей после переноса различных клеточных популяций. Введение нефракционированных Т-клеток CD4+предотвращало развитие колита, тогда как фракция СD4-клеток, экспрессирующих высокий уровень CD5RB (CD45RBhl), вызывала колит. При одновременном введении антител анти-ИФγ или анти-ФНОα частота случаев заболевания снижалась. Цитокин ИЛ-10 (но не ИЛ-4) также влияет на возникновение колита у мышей. Представленные данные показывают, что в патогенезе колита у мышей играют роль цитокины ИФα и ФНОα и что ИЛ-10 может инактивировать эффекторные клетки. (ТКИД - тяжелый комбинированный иммунодефицит.)