Эволюция вирусов - Жданов В. М. 1990
Специальная часть
РНК-содержащие изометрические вирусы
В этой главе будут рассмотрены РНК-содержащие вирусы с икосаэдральным строением вирионов, которые представляют сборную группу, имеющую, несомненно, и разное происхождение, и разные пути эволюции. Несмотря на давнее «знакомство» с большинством из них, они изучены недостаточно. Все они имеют положительно-полярный геном, молекулярная масса которого колеблется в пределах от 1,2 x 106 до 3,2 х 106, диаметр вирионов составляет 23—35 нм [Matthews R., 1982].
Наиболее мелкими являются вирусы семейства Leviviridae, поражающие разные виды бактерий. Их геном имеет молекулярную массу 1,2 х 106, составляя 30% массы вирионов, на нем закодированы 4 белка: капсидный (12000—14000), белок созревания (35000—44000), репликаза и литический белок. Капсидных белков на вирион приходится 180 молекул, белка созревания — одна молекула. Репликаза синтезируется в ходе инфекции. Вирионы являются икосаэдрами с диаметром 23 нм, состоят из 32 капсомеров. Эта группа фагов насчитывает около 40 вирусов, имеет узкую специфичность — они поражают мужские особи энтеро и каулобактерий, псевдомонад.
Вирусы группы Nudaurelia ß (6 вирусов) поражают насекомых (разные виды чешуекрылых). Молекулярная масса РНК составляет 1,8 x 106, т. е. 10—11% массы вирионов. Вирионы содержат один капсидный белок (60 000—70 000) - 240 субъединиц (Т-4), имеют диаметр 35 нм. Большинство вирусов этой группы иммунологически родственны.
У насекомого Nudaurelia cytherea capensis обнаружено 6 разных вирусов. Из них более подробно описан вирус Сu. Имеющий белок с молекулярной массой 65 000 [Hendry D. et al., 1985].
Среди вирусов растений сходны вирусы трех групп. Первую группу составляет вирус хлоротической карликовости маиса (MCDV) и сходный вирус риса тунгро. Геном имеет молекулярную массу 3,2 x 106, белки не изучены. Вирионы представляют собой полиэдры диаметром 30 нм. Передаются тлями. Род Luteovirus представлен вирусом желтой карликовости ячменя (BYDY) и другими (около 40) вирусами растений. Их геном имеет молекулярную массу 2,0 x 106, а белок капсида 24 000. Вирионы изометрические, диаметром 25— 30 нм. Реплицируются во флоэме растений. Передаются тлями, у которых вирусы персистируют. Спектр «хозяйской» специфичности варьирует у разных представителей данного рода, значительная часть которых отнесена к нему условно.
Род Tombusvirus включает около 11 вирусов, поражающих растения, в том числе вирус кустистой карликовости томатов (TBSV). Они имеют небольшой геном с молекулярной массой около 1,5 х 106, составляющий 17% массы вирионов, один капсидный белок (41000). В вирионе содержится 180 молекул капсидного белка, они образуют округлые частицы диаметром 30 нм. Большинство вирусов этого рода перекрестно иммуногенны. Репликация их сходна с репликацией калицивирусов: в ходе ее образуется наряду с полным двунитевым геномом субгеномная РНК. Вирусы поражают широкий круг растений, передаются механически.
В группе томбусвирусов имеется 6 серологически родственных вирусов, у большинства обнаружена значительная гомология нуклеиновых кислот, несмотря на то что размеры генома варьируют от 3500 (galinsoga mosaic virus) до 4700 (tomato bushy stunt virus) нуклеотидов. Однако наиболее близкие по антигенам вирусы, образующие группу, имеют геном размером в 4700 нуклеотидов [Gallitelli D. et al., 1985]. С помощью теста двойной диффузии в агаровом геле были определены индексы серологической дифференциации томбусвирусов, на основании которой была составлена дендрограмма серологического родства между 10 томбусвирусами (рис. 7). Эти данные коррелируют с аминокислотными последовательностями и могут явиться основанием для предположений об эволюции томбусвирусов [Koenig R., Gibbs А., 1986].
Уместно рассмотреть еще несколько групп вирусов с позитивно-полярным геномом.
Несколько (6) вирусов, поражающих насекомых, также являются бисегментными, образуют семейство Nodaviridae. Их геном состоит из двух сегментов РНК с молекулярной массой 1,15 x 106и 0,5 х 106. Эти сегменты расположены в одной частице диаметром 29 нм. РНК не содержит поли(А) - последовательностей. Первый сегмент кодирует белок с молекулярной массой 105 000 (вероятно, репликазу), второй — белок-предшественник (43 000) капсидного белка. Мажорный белок имеет молекулярную массу 40 000, минорный — 43 000 (или два по 39 000). Кроме того, в зараженных клетках образуются низкомолекулярные пептиды с молекулярной массой 10 000 и 5000. Первый участвует в репликации, а второй входит в состав вирионов. Вирусы поражают двукрылых, чешуекрылых, перепончатокрылых, круг хозяев обычно широк.
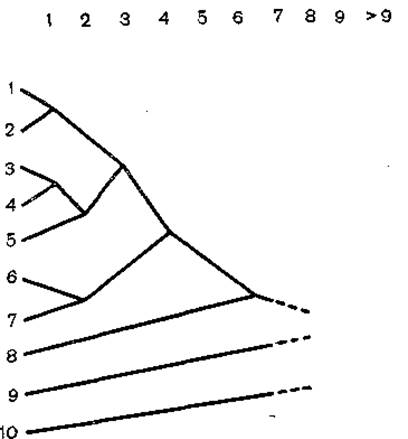
Рис. 7. Дендограмма, иллюстрирующая взаимосвязь 10 томбусвирусов по реципрокньм тестам.
1 — TBSV; 2— TBS K-BS3; 3 — АМС V; 4 — PAMV; 5 — PLCV; 6 — MPV; 7 — EMCV; 8 — CIRV; 9 — TNV; 10 — Cyb RSV.
Род Dianthouirus представлен вирусом кольцевой пятнистой гвоздики и еще двумя вирусами. Геном их бипартитлый с сегментами РНК 1,5 x 106 и 0,5 х 106, причем в первом находится ген капсидного белка (40 000). Частицы являются полиэдрами диаметром 31—34 нм. Круг хозяев широк, вирусы передаются механически.
Группа pea enation mosaic virus пока включает одного представителя, который является бипартитным вирусом. Его РНК1 имеет молекулярную массу 1,77 x 106 и кодирует два мажорных белка Р2 (88 000) и Р4 (369 000), один минорный белок Р1 (147000). Гены этих белков расположены в следующем порядке: Р4 — Р2 — P1. РНК2 имеет молекулярную массу 1,2 x 106 и кодирует белок Р3 (45 000). Капсидным белком является VP2 [Gabriel С., De Zolten С., 1984].
Группа вирусов бархатной мозаики табака (velvet tobacco mottle virus group) насчитывает 4 представителя, имеет два фрагмента РНК (1,5 x 106 и 0,12 x 106), из которых второй — циркулярный. Оба фрагмента находятся в разных частицах диаметром 30 нм. Цикл репродукции сложный, включает ядерные и цитоплазматические синтезы. Вирусы передаются жуками и миридами (myrids), а также механически.
Вирус мозаики гороха (enation mosaic) выделен в самостоятельную группу. Его геном также бипартитный с сегментами РНК 1,7 х 106 и 1,3 х 106. Имеется два капсидных белка — мажорный (22 000) и минорный (28 000). РНК заключена в полиэдральные частицы диаметром 28 нм. Вирус передается тлями (aphids), в организме которых он персистирует.
Вирус рода I lаr является бипартитным. Он имеет 4 РНК с молекулярной массой 1,1 х 106; 0,9 х 106; 0,7 x 106, 4-я РНК — это мРНК (0,3 x 106) для синтеза капсидного белка (25 000). Частицы для разных РНК имеют диаметр 26—35 нм. Цикл репродукции довольно сложный. Пятнадцать вирусов поражают широкий круг растений и передаются либо механически, либо семенами.
Трипартитный вирус мозаики люцерны имеет три РНК, которые кодируют белки с молекулярной массой 126000 (РНКІ), 90 000 (РНК2), 32 000 (РНК3); 4-й компонент — это мРНК (РНК4), гомологичная 3'-терминальной последовательности РНКЗ. Первые два белка обеспечивают синтез вирусных РНК, в то время как третий белок регулирует баланс между плюс- и минус-нитями вирусной РНК/ РНК4 кодирует синтез капсидного белка [Sarachu A. et al., 1985].
Вкратце напомним еще о трех группах изометрических РНК-содержащих вирусов, поражающих растения. Одна из них, род Necrovirus, включает в себя вирусы некроза табака и огурцов. РНК (1,3 x 106, 1,6 x 106) имеет на 5'-конце структуру ppApGpUp. Белок капсида один (22 000). Вирионы представляют собой полиэдры диаметром 28 нм, поражают ангиоспермы, передаются грибом Olpidium. Вторая группа — род Tymovirus — представлена вирусом желтой мозаики турнепса и еще 17 вирусами. РНК (2 х 106) составляет 35% массы вирно-нов и имеет на 5'-конце кэп-структуру m7G5'ppp5'Gp и структуру, сходную с тРНК, на 3'-конце. В некоторых частицах обнаруживается мРНК (0,2x10s—0,3x10s) для синтеза капсидного белка — 180 молекул на вирион. Из двух типов частиц ß-частицы содержат полный геном, а T-частицы являются продуктом несбалансированных синтезов. Морфологически это икосаэдры диаметром 29 нм, состоящие из 20 гексамеров и 12 пентамеров. Среди представителей рода имеется иммунологическое родство. Вирусы поражают двудольные (Licotyledonous) растения, передаются механически или жуками. Вирусы третьей группы — вирусы мозаики огурцов (род Cucumovirus) — насчитывают 4 представителя, являются трипартитными: 1,25 x 106 (РНК1); 1,13 х 106 (РНК2); 0,82 x 106 (РНКЗ). Четвертый тип частиц содержит мРНК ((0,35 x 106) для синтеза белка капсида. В отличие от РНК предыдущей группы вирусов все 5'-концы РНК имеют кэп-структуру m7G5'ppp5'Np, а на 3'-конце — тРНК-подобную структуру, акцептирующую тирозин. Фрагменты РНК заключены в икосаэдрические частицы диаметром 29 нм. Каждая из них транслируется в отдельный белок, причем первые три кодируют белки с молекулярной массой 105 000, 120 000 и 34 000 [Афанасьев Б. Н. и др., 1986]. Вирусы передаются механически, через семена и тлями.
Сопоставленные в этой главе вирусы имеют как общие, так и различающие их на подгруппы черты. Общими чертами являются позитивная полярность РНК, отсутствие 5'-терминальных белков, изометрический тип строения вирионов (или части). Далее идут различия.
В первых 5 группах пока не обнаружен механизм разделения синтеза продуктов отдельных генов, хотя на примере мелких изометрических фагов показано, что активность каждого из 4 генов регулируется в процессе трансляции. Так, синтез полимеразы прекращает активность гена, ее кодирующего, в связи с блокировкой исходной точки считывания. Вероятно, по этому типу регулируется деятельность немногих генов, которые имеются у остальных трех групп вирусов. Экологически ниши их разнообразны: бактерии, насекомые, растения (передача тлями с персистенцией в них вирусов, а также грибами).
У томбуc и тимовирусов уже имеется своеобразный механизм разделения активности разных генов — синтез субгеномной РНК, несмотря на то что этот же участок той же позитивной полярности есть на геномной РНК. Синтез субгеномных РНК повторится у калицивирусов, геном которых также имеет позитивную полярность, но отличается наличием белка, ковалентно связанного с его 5'-концом.
Мультипартитные вирусы можно рассматривать как дальнейшее развитие разделения функционирования отдельных генов и поэтому в третью группировку помещены вирусы с разобщенным геномом. Сюда относятся, во-первых, нодавирусы, поражающие насекомых, у этих вирусов геном разделен на два фрагмента, хотя оба фрагмента находятся в одном и том же капсиде; большой фрагмент кодирует полимеразу, а малый — капсидный белок. Три группы вирусов растений с бипартитнымй геномами содержат полный геном в разных частицах; вирус мозаики гороха может содержать 3-й компонент РНК. Группы трипартитных вирусов растений могут образовывать 4-й компонент — мРНК для синтеза капсидного белка. Все или почти все гены механически (пространственно) разобщены.
Таким образом, рассмотренные группы мелких РНК-содержащих вирусов иллюстрируют одно из возможных направлений эволюции их генома, обеспечивающее раздельный синтез кодируемых разными генами белков, количественные соотношения которых, конечно же, не должны быть эквимолярными, С этих позиций функционирование небольших геномов, содержащих не больше 4—5 генов, легче регулируется, если они разобщены, а это почти идеально происходит в мультипартитных вирусах. Но мультипартитность имеет и «обратную сторону»: для эффективного заражения необходимо одновременное попадание в организм нового хозяина всех компонентов мультипартитного вируса. Указанные группы вирусов не представляются связанными между собой, а некоторые из них, например левивирусы, являются узко специализированными группами, поражающими только мужские особи некоторых видов бактерий. Две группы вирусов поражают насекомых. Из остальных 7 групп вирусов, поражающих растения, 4 группы передаются насекомыми, преимущественно тлями. Последнее может свидетельствовать о возможном способе освоения этими вирусами новых экологических ниш.
Имеются, однако, основания предполагать, что разные группы вирусов растений имеют эволюционные связи. В частности, это было показано при сравнительном изучении аминокислотного состава бромовирусов (трехпартитные, кэп-тРНК-структуры), кукумовирусов (трехпартитные, кэп-тРНК-структуры), вируса мозаики люцерны (четырехпартитный, кэп-структуры). Дендрограмма, приведенная на рис. 8, показывает возможную дивергенцию этих веществ от единого предка [Dale J. et al., 1985].
Вероятно, этими соображениями можно и ограничиться, анализируя возможные источники происхождения рассматриваемых групп вирусов. Эволюционные связи между разными вирусами, имеющими между собой иммунологическое родство, вполне возможно, выявятся не только при секвенировании геномов, но и при сопоставлении возможных путей эволюции вирусов и эволюции их хозяев. Например, при анализе вирусов группы Nudaurelia ß явно нужно изучение территориального распространения и филогенеза их хозяев — чешуекрылых. Весьма полезными могут оказаться филогенетические соотношения между энтеробактериями, родами Саulobacter и Pseudomonas, а также экологические взаимоотношения между Escherichia и Podellvibrio при анализе изометрических РНК-содержащих фагов. Интересно сопоставление серологического родства между вирусами группы желтой карликовости ячменя с историей культивирования злаковых, моркови и турнепса, гороха, сои и клевера и других культурных растений, а также «спектром» паразитизма тлей, поражающих эти растения.

Рис. 8. Дендограмма, иллюстрирующая классификацию белков бромовирусов, кукумовирусов и вируса птичьего миелобластоза, аминокислотный состав которых получен с помощью компьютера.
1 — ВВМ 1; 2 — ВВМ 2; 3 — BBM 3; 4 — ССМ 1; 5 — ССМ 2; 6 — ССМ 3; 7 — ВМ; 8 — CYB; 9 — AMVS- 10 — AMV-425; 11, 12 — вирус мозаики огурцов 1 и 2; 13 — PS; 14 — вирус аспермии томатов.
При рассмотрении мелких изометрических РНК-содержащих вирусов мы исходили из того, что они не имеют ни кэп-структур, ни 5'-терминальных белков. Дальнейшее изучение их покажет, является ли это положение справедливым для всех рассматриваемых групп вирусов. Однако специальное указание на две важные особенности репликации РНК вирусов, возможно, будет способствовать большему пониманию излагаемого материала. Конкретно к таким особенностям можно отнести следующие. 1. При РНК-зависимом синтезе РНК отсутствует механизм исправления ошибок репликации (частота «опечаток» может достигать 10-3 против 10-7—10-10 для ДНК). 2. Отсутствие ферментативного аппарата для разрушения протяженных двунитевых структур РНК. В результате изменяется стратегия репликации генома, появляются специальные механизмы и структуры, предотвращающие образование протяженных дуплексов РНК при репликации (тРНК-подобные концевые структуры, одновременное взаимодействие сразу с несколькими молекулами репликазы, трансляция растущих плюс-цепей РНК и т. д.).
Гомология между вирусными и клеточными белками отмечается и у вирусов прокариотов [Koji О., 1985]. Так, ß-субъединица репликазы фага MS2 имеет гомологию с а-субъединицей РНК-полимеразы кишечной палочки, есть гомология между С-концом упомянутой выше вирусной полимеразы, ДНК-примазой (продукт гена dnaG) и а-субъединицей полимеразы кишечной палочки, имеется гомология между ß-peп-ликазой фага MS2, ДНК-полимеразой фага Т7 и N-концом а-субъединицы бактериальной полимеразы. Эти примеры позволяют сделать вывод о том, что эволюция идет блоками (модулями), которые могут использоваться возникающими из клеточных элементов вирусами, иногда весьма различными (РНК-содержащий фаг MS2 и ДНК-содержащий фаг 77), либо же речь может снова идти о молекулярной конвергенции. Дополнительное изложение этих особенностей репликации в главах, посвященных описанию структуры РНК-содержащих вирусов, может быть, позволит в большей мере понять появление фрагментированных геномов, необходимость компактизации генетической информации и др.
Наконец, следует отметить некоторые особенности экономного использования генома (ниже приводятся такие механизмы у ДНК-содержащих вирусов): 1) сдвиг рамки структурных генов (MS2, J2 РНК, ФХ174 ДНК); 2) разные стоп-сигналы при одинаковом начале транскрипции (Qß, TYMV, РНК, ФХ174 ДНК); 3) транскрипционные сигналы (JD, РНК, ФХ174 ДНК); 4) разные направления считывания (фаг Я). Более подробные комментарии [Reanney D., 1984] относятся к РНК-содержащим вирусам.