ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 1 - Г.Й. Щербак - 1995
ПІДЦАРСТВО НАЙПРОСТІШІ, АБО ОДНОКЛІТИННІ (PROTOZOA)
ТИП ВІЙКОНОСНІ, АБО ІНФУЗОРІЇ (CILIOPHORA)
Представники типу мешкають переважно у водному середовищі, серед них є також симбіотичні та паразитичні форми. Відомо близько 7,5 тис. видів інфузорій. Це організми порівняно великих розмірів (50—300 мкм); окремі види (1—3 мм) вкриті війками протягом усього життя або лише на певних фазах життєвого циклу. Для них характерний ядерний дуалізм — одне вегетативне ядро великих розмірів (макронуклеус) і одне або кілька генеративних ядер (мікронуклеус). Інфузорії розмножуються поділом або брунькуванням, а також мають особливий тип статевого процесу — кон'югацію.
Ектоплазма (кортекс) інфузорій має дуже складну будову, яку вивчено за допомогою електронно-мікроскопічних методів. У кортексі містяться різноманітні структури, що забезпечують сталість форми тіла. Зовнішня частина кортексу — пелікула — утворена плазматичною мембраною (плазмалемою) та розташованими під нею сплющеними мішечками, які мозаїчно з'єднані між собою. У деяких видів всередині мішечків локалізовані додаткові, опорні структури — білкові або просякнуті вуглекислим кальцієм полісахаридні пластинки. Під пелікулою розташовані базальні тільця (кінетосоми) всіх війок, а також, зв'язані з ними утвори. У більшості інфузорій це три постійні компоненти, які відходять від кінетосоми: 1) розташований під пелікулою поперечносмугастий філамент, що спрямований до переднього кінця клітини; 2) група мїкротрубочок, спрямованих уздовж тіла до його заднього кінця; 3) група мікротрубочок, розташована поперек тіла. Сукупність цих трубочок утворює єдиний цитоскелет, що міститься під пелікулою. Цитоскелет — ектоплазматична фібрилярна система є лише в інфузорій (рис. 44).
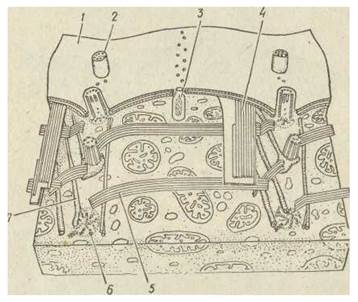
Рис. 44. Ультраструктура кортексу в Ciliata
1 — пелікула; 2 — війки; 3 — трихоциста; 4 — поздовжні мікротрубочки;
5 — поперечні мікротрубочкн; 6 — кінетосома; 7 — мікрофіламент
У пелікулі перпендикулярно до її поверхні містяться екструсоми — пухирцевидні утвори, всередині яких міститься особливий білок. Вони вистрілюються клітиною під час подразнення, багаторазово подовжуючись внаслідок розтягування білкових молекул. У більшості війчастих вони мають вигляд видовжених тілець, добре помітних під світловим мікроскопом і називаються трихоцистами. Викидаються трихоцисти назовні під час будь-якого сильного механічного чи хімічного подразнення, висихання тощо, їх функцію остаточно не з'ясовано. Припускається, що вони містять отруйні речовини, які вбивають ворогів або виконують функції осморегуляції, прикріплення тощо.
Класичний об'єкт лабораторних досліджень — інфузорія туфелька (Paramecium caudatum) — має 5—8 тис. трихоцист. У хижих інфузорій є інші екструсоми — токсоцисти. Вони мають вигляд капсули з оболонкою з трубочок, у порожнині якої міститься внутрішня трубка. Під час полювання ця трубка вивертається назовні, як палець рукавички, або телескопічне висувається з капсули, пронизує тіло здобичі (інші найпростіші, коловертки тощо) та впорскує в неї отруту, паралізуючи, чи вбиваючи.
Війки інфузорій — органели руху — за тонкою будовою не відрізняються від джгутиків. У найпростіших інфузорій вони рівномірно вкривають усю поверхню тіла, проте часто концентруються на певних ділянках або спеціалізуються, утворюючи циррі, мембрани або мембранели. Циррі — це пучечки або китиці з війок, завдяки яким найпростіше може «бігати» по субстрату чи «стрибати» в товщі води.
Мембрани — це ряди війок, з'єднані між собою. У мембранелах війки розташовані в ряд, однак не сполучені одна з одною. Мембрани та мембранели забезпечують рух води до ротового отвору. Сукупність усіх війок та їх похідних називається ціліатурою.
Війки рухаються узгоджено. Веслоподібний рух війок складається з двох фаз — робочий удар і зворотний рух. Під час робочого удару війка згинається лише біля основи, залишаючись прямою й женучи рідину в напрямку свого руху. Під час зворотного руху вона повністю згинається, стає м'якою й проходить поблизу поверхні тіла, майже не захоплюючи воду — так здійснюється поступальний рух. Війки розташовані рядами, що називаються кінетами. В кожній кінеті сусідні війки перебувають у одній фазі биття, однак порівняно з сусідньою кінетою їх рух завжди трохи зсунутий за фазою, що низавається метахронією. Наприклад, у туфельки ряди війок, що перебувають у одній фазі руху, розташовані навскоси щодо поздовжньої осі тіла — під мікроскопом видно, що по поверхні клітини ззаду наперед неначе пробігають хвилі. Причина такого узгодження полягає не в структурі цитоплазми, а в гідродинамічних властивостях самих війок, тобто течія води, створювана однією війкою, синхронізує фази руху сусідніх.
Більшість інфузорій, крім деяких ендопаразитичних форм (ряд Astomatida), має клітинний рот — цитостом, Найпростішою формою ротового апарата є термінальний
рот, що міститься на передньому кінці тіла й оточений звичайною соматичною ціліатурою (Holophrya, Prorodon). Рот більшості інфузорій розміщений на бічній частині тіла й заглиблений у вп'ячування, або перистом (вестибулюм), на дні якого відкривається клітинний рот. У деяких видів перистом веде до вузького каналу — глотки, або цитофаринксу, що закінчується в ендоплазмі (рис. 45, 46).
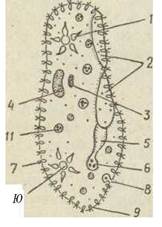
Рис. 45. Схема будови Paramecium caudatum:
1 — передня скоротлива вакуоля; 2 — навколоротове заглиблення (перистом); 3 — мікронуклеус; 4 — макронуклеус; 5 — глотка (цитофаринкс); 6 — клітинний рот (цитостом); 7 — війки; 8 — порошиця; 9 — трихоциста; 10 — задня скоротлива вакуоля; 11 — травна вакуоля
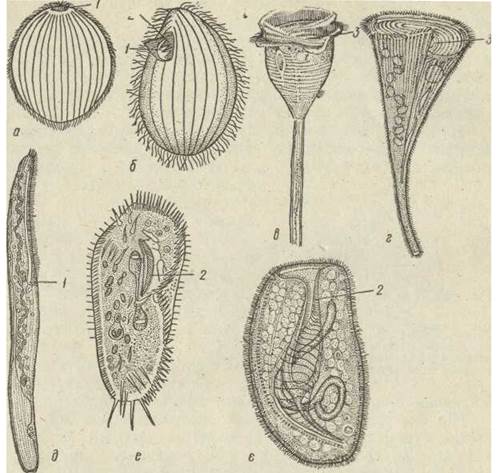
Рис. 46. Цитостом і ціліатура в інфузорій — Holophrya (a); Tetrahymena pyriformis (б); Vorticella sp. (в); Stentor polymorphys (г); Spirostomum ambiguum (д); Stylonichia mytilus (e); Bursaria truncateHa (є):
1 — цитостом; 2 — мембранели; 3 — перистом
Ціліатура навколо ротового апарата теж диференціюється, війки зливаються у мембранели, що призначені для спрямовування їжі до рота. У найпростіших інфузорій — це три паралельно розташовані мембранели з одного боку рота й одна ундулююча мембрана — з іншого. Мембранели створюють течію води, з якою частки їжі надходять до ундулюючої мембрани, яка спрямовує їх до рота. Цей комплекс органел називається тетрахіменіумом (від назви інфузорії Tetrahymena, що має такий апарат у найбільш типовому вигляді). Навколоротову ціліатуру мають також інфузорія туфелька (Paramecium caudatum) і, в дещо видозміненому вигляді — сидяча сувійка (Vorticella marginata).
Навколоротовий апарат складнішої будови — це спірально закручена праворуч зона навколоротових мембранел, що інтенсивно заганяють їжу до рота. Таку ротову ціліатуру мають інфузорії, які часто трапляються в прісних водоймах: Stentor, Spirostomum, Stylonichia тощо. Інфузорії з описаною навколоротовою ціліатурою живляться, підганяючи за її допомогою дрібні часточки їжі до рота й заковтуючи їх. Форми, що не мають навколоротової ціліатури, є переважно хижаками. Вони живляться іншими найпростішими, заковтуючи їх (Didinium). Деякі заковтують тонкі нитчасті водорості (Pseudomicrothorax dubius, Nassula). У цих інфузорій є складне пристосування до заковтування їжі — паличковий апарат. Він складається з сотень мікротрубочок, зібраних у правильні пучки й пластинки, які утворюють трубку, що міститься в цитоплазмі навколо рота та виконує опорну функцію під час заковтування цілої здобичі (рис. 47).
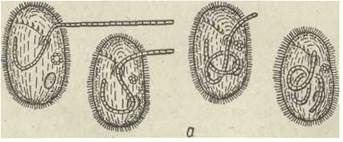
Рис. 47. Nassula sp.:
a — стадії заковтування ниток синьо-зелених водоростей; б — паличковий апарат
Сисні інфузорії (Suctoria) цитостому не мають, у них є багато сисних щупалець. В кожному з них розвинена трубка, що складається з пучків мікротрубочок. За її допомогою цитоплазма здобичі, що прилипає до такого щупальця, всмоктується в тіло сисної інфузорії.
Найбільш детально процеси живлення та травлення досліджені у Paramecium caudatum. Перистом у неї переходить у цитофаринкс, який закінчується в ендоплазмі цитостомом — сліпою кишенею, відокремленою від ендоплазми лише однією мембраною. В цій кишені накопичуються часточки їжі й формується травна вакуоля, яка відшнуровується, а на її місці утворюється нова кишеня. Травна вакуоля переміщується в тілі інфузорії, проходячи шлях від місця утворення через усе тіло, й закінчує своє існування в клітинній порошиці, або цитопрокті.
Під час руху вакуолі в ній перетравлюється їжа. Цей процес супроводжується зміною pH середовища всередині вакуолі. До травної вакуолі транспортуються дрібні пухирці, які містять кислоту й викликають різку зміну pH (від нейтрального до кислого), що необхідно для стимуляції дії травних ферментів. Потім до вакуолі підходять і зливаються з нею лізосоми, що несуть травні ферменти. Після перетравлення їжі дрібні пухирці з поживними речовинами відшнуровуються від травної вакуолі й розподіляються по клітині, а сама вакуоля, в якій залишилися неперетравлені рештки, підходить до спеціального (обмеженого лише мембраною) отвору в пелікулі — порошиці, що розташована недалеко від перистома. Після викиду решток їжі через порошицю вакуоля розпадається на пухирці, які повертаються до цитостому й входять до складу нових травних вакуолей. Травлення у інфузорій триває від 20 хв до кількох годин (рис. 48).

Рис. 48. Травлення в Paramecium:
1 — утворення травної вакуолі в цитостомі; 2—5 — різні стадії циклу травної вакуолі; 6 — травна вакуоля з рештками їжі;
7 — порошиця, через яку рештки їжі викидаються
Скоротливі вакуолі інфузорій, яких у однієї особини міститься одна або кілька, мають складну будову. Вакуоля — це скоротливий пухирець, що відкривається отвором (порою) назовні. В неї впадають зірчасте розташовані ампули, тобто кінці довгого тонкого каналу-провідника. Міцність та фіксоване положення цього комплексу в тілі зумовлюють стрічки з мікротрубочок, що починаються від стінок видільної пори та йдуть уздовж ампул і каналів. Ці стрічки в свою чергу укріплені спіральне розташованими мікротрубочками (рис. 49).
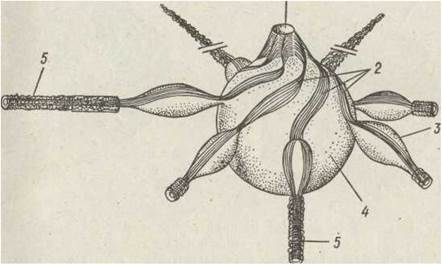
Рис. 49. Схема будови скоротливої вакуолі Paramecium:
1— пора; 2 — стрічки мікротрубочок; 3 — ампули; 4 — скоротлива вакуоля; 5 — провідні канали
Для багатьох війчастих на певних етапах життєвого циклу характерний розвиток прикріпних апаратів. Наприклад, у Vorticella прикріпне стебельце утворюється особливим органоїдом — скопулою. Це чашеподібна, оточена валиком заглибина на задньому (протилежному від рота)
полюсі клітини, де містяться численні нерухомі війки. Цей утвір може видовжуватися тa перетворюватися на стебельце. У ектопаразита риб Trichodina є спеціальний присосок, побудований з періодично повторюваних білкових структур (рис. 50).
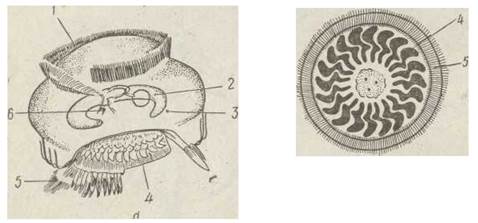
Рис. 50. Схема будови (о) та прикріпний апарат (б) інфузорій роду Trichodina:
1 — навколоротова ціліатура; 2 — макронуклеус; 3 — мікронухлеус;
4 — прикріпний апарат; 5— базальний віночок війок; 6 — цитостом
Нестатеве розмноження інфузорій має характер поділу. Під час поділу мікронуклеус ділиться мітотичне, макронуклеус перешнуровується навпіл. Невідомо, як при цьому розподіляються хромосоми в дочірніх ядрах — випадково (амітоз) чи якимось іншим чином. У процесі поділу цитоплазми, як правило, поперек тіла утворюються по два нові перистоми, рота, глотки, деякі органоїди розподіляються між дочірніми клітинами з відновленням тих структур, яких не вистачає. В ендоплазмі утворюються кінетосоми, що мігрують до плазмалеми, де з них розвиваються війки.
Якщо поділ клітини нерівномірний, він має характер брунькування. Брунькування характерне для сидячих форм, від яких відбруньковуються розселювані стадії — бродяжки. У сисних інфузорій такі бродяжки утворюються в спеціальній камері всередині тіла, причому з материнським організмом їх з'єднує стебельце — своєрідний аналог плаценти, а потім через спеціальний отвір виходять назовні.
Під час статевого процесу інфузорії злипаються бічними поверхнями, в області цитостому в них виникає цитоплазматичний місток. Статевий процес (кон'югація) не супроводжується утворенням гамет, замість них зливаються гаплоїдні ядра. Цікаво, що особини одного й того виду кон'югують тільки з особинами, які належать до комплементарного типу парування. Таких типів у Р. caudatum налічується 40, причому паруватися вони можуть лише в певних комбінаціях. Інфузорії, що належать до одного типу, не кон'югують між собою. Інфузорії комплементарних типів парування пізнають одна одну за допомогою спеціальних біологічно активних речовин — гомонів, що виділяються ними в зовнішнє середовище. Ці речовини змінюють властивості поверхні клітини комплементарної особини, зумовлюючи кон'югацію.
Кон'югація в різних видів різниться деталями, однак відбувається за загальною схемою. У туфельки цей процес починається з того, що мікронуклеус збільшується, а макронуклеус розсмоктується. Мікронуклеус ділиться мейотичне, з чотирьох гаплоїдних ядер, що утворилися, три розсмоктуються, четверте мітотичне поділяється на стаціонарне (жіноче) та мігруюче (чоловіче) ядра — пронуклеуси. Партнери обмінюються мігруючими ядрами, кожне з яких зливається зі стаціонарним ядром іншої особини з утворенням диплоїдного ядра (синкаріон), після чого особини розходяться.
Потім синкаріон шляхом послідовних мітозів утворює вісім ядер, з яких три гинуть, одне стає мікронуклеусом, а чотири — зачатками макронуклеусів. Внаслідок поділу мікронуклеуса ці протоплазми утворюють дві клітини з одним мікронуклеусом і двома зачатками макронуклеусів (цей процес повторюється). Зрештою, утворюються чотири клітини, кожна з яких містить по одному мікронуклеуйу та одному зачатку макронуклеуса (обидва ядра диплоїдні). У зачатку макронуклеуса більшість хромосом гине, а ті, що залишилися, розпадаються на дрібні ділянки—окремі гени, більша частина яких також руйнується. Потім починається інтенсивний синтез ДНК, що веде до збільшення числа генів, що залишилися, в сотні та тисячі разів. Отже, хоча макронуклеус поліплоїдний, в ньому залишається лише невелика частина (1,6 %) генома мікронуклеуса (рис. 51.)
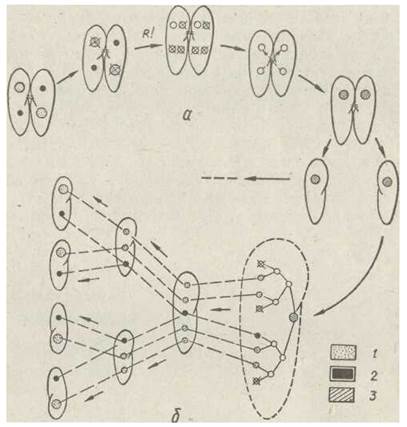
Рис. 51. Послідовні стадії змін ядерного апарата (а) та його відновлення (б) у Paramecium caudatum у процесі кон’югації:
1 — макронуклеус; 2 — мікронуклеус; 3 — синкаріон; R! — редукційний поділ
Цикл розвитку інфузорій проходить з гаметичною редукцією. Його особливості — відсутність гамет і ядерний дуалізм. Якщо інфузорія не має партнера для кон'югації, статевий процес відбувається автогамно, тобто мігруюче та стаціонарне ядра зливаються всередині однієї особини. Біологічний зміст кон'югації — обмін спадковим матеріалом між особинами, що веде до комбінаційної мінливості. Істотне значення має також відновлення макронуклеуса: під час поділів кількість генів у ньому зменшується, що сповільнює розвиток і ріст інфузорій.
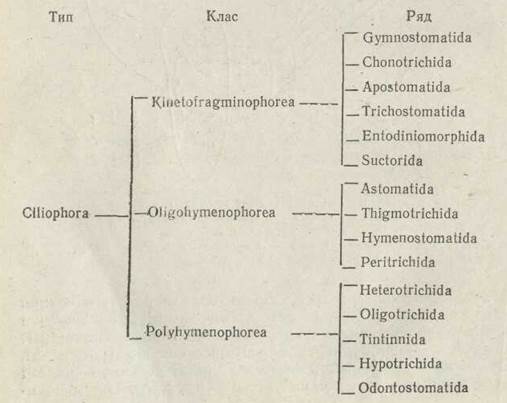
Система інфузорій за останні роки зазнала численних переробок, пов'язаних з новими електронно-мікроскопічними дослідженнями, які поглибили знання про їх будову. Було запропоновано кілька варіантів класифікації інфузорій, але єдиної, загальновизнаної системи досі ще не існує. На сьогодні найбільш поширеною є класифікація, в основу якої покладено організацію ротової циліатури інфузорій. За цією системою тип поділяється на три класи: