ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 1 - Г.Й. Щербак - 1995
ПІДЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
ЗАГАЛЬНА ХАРАКТЕРИСТИКА
Багатоклітинні характеризуються. тим, що їх тіло складається з багатьох клітин та їх похідних. Проте на відміну від колоній найпростіших, до складу яких також входить велика кількість клітин, у тілі Metazoa клітини диференційовані й виконують різні функції. Вони втрачають свою індивідуальність, стають частинами складного організму, й, об'єднуючись для виконання певних функцій у більшості .багатоклітинних, утворюють тканини: епітеліальну, ,сполучну, м'язову, нервову. Тканини в свою чергу утворюють складні органи та системи органів, які працюють в організмі узгоджено, й він функціонує як єдине ціле. Отже, характерною ознакою Metazoa є поява спеціалізованих тканин, органів і систем органів.
Особливістю багатоклітинних на відміну від колоній найпростіших е багатошарове розташування їх клітин, за якого зовнішні клітини утворюють суцільний ЕІар, що відокремлює тіло тварини від зовнішнього середовища. В такий спосіб у багатоклітинних виникає внутрішнє середовище організму, де містяться всі клітини його тіла й підтримується постійність фізико-хімічних параметрів.
Багатоклітинним тваринам властиве нестатеве та статеве розмноження, проте переважаючою формою, а в деяких групах і єдиною, є статеве.
Нестатеве (вегетативне) розмноження багатоклітинних— це форма розмноження без участі статевих клітин. Воно може відбуватися двома способами: поділом (поперечним, поздовжнім або неупорядкрваним) та брунькуванням (внутрішнім або зовнішнім).
Усі багатоклітинні розмножуються статевим способом (інколи партеногенетично). Статеве розмноження у багатоклітинних здійснюється за допомогою спеціальних генеративних (статевих) клітин. Решта клітин — соматичні. Соматичні клітини у них дишіоїдні, а гамети, що утворюються з генеративних клітин — гаплоїдні — гаметична редукція (див. рис. 22). Характерною особливістю статевого процесу багатоклітинних є оогамія: яйцеклітини (макрогамети) та сперматозоїди (мікрогамети) різняться не лише за розмірами, а й за структурою. Сперматозоїд у типовому випадку має будову дещо видозміненого джгутиконосця. Нерухомий сперматозоїд має назву спермію (нематоди, десятиногі раки тощо). Яйцеклітини (яйця) нерухомі, позбавлені джгутиків і мають переважно сферичну форму. Деякі з них рухаються амебоїдно (наприклад, у гідри). В цитоплазмі яйцеклітин більшості тварин містяться жовточні гранули — запаси поживних речовин (білків,, полісахаридів, жирів), а також нуклеїнові кислоти, ферменти. Лише у нижчих груп яйця не мають жовтка. В яйцеклітині розрізняють два полюси: анімальний, де містяться ядро та зона інтенсивного обміну речовин, і протилежний — вегетативний.
Яйце оточується однією або кількома оболонками. Склад і будова цих утворів у різних тварин неоднакові.
Особливою формою розмноження, яка виникла на базі статевого розмноження, є партеногенез, за якого новий організм утворюється з незаплідненого яйця.
Життєвий цикл в усіх Metazoa характеризується складним індивідуальним розвитком — онтогенезом, в процесі якого із заплідненого яйця утворюється дорослий організм.
Онтогенез багатоклітинних складається з ряду етапів. Спочатку утворюються гамети (гаметогенез). При цьому недиференційовані диплоїдні клітини мейотично діляться й утворюються гаплоїдні яйцеклітини та сперматозоїди. Після запліднення починається дробіння яйця, яке має характер палінтомічного поділу. Дробіння завершується утворенням одношарового зародка — бластули. Після закінчення дробіння клітина ділиться монотомічно, починаються процеси диференціації та переміщення клітин (гаструляція), які спочатку приводять до утворення двох або трьох зародкових листків (ектодерми, ентодерми та мезодерми), а пізніше — зачатків органів. Від цієї схеми відрізняється онтогенез первинних багатоклітинних (Prometazoa), які не мають зародкових листків. Значна частина онтогенезу (ембріональний розвиток) проходить під покривом яйцевих оболонок (або в тілі материнського організму). Після виходу з яйця (або народження) починається постембріональний розвиток, який буває прямим (коли молодий організм- подібний до дорослого) або супроводжується метаморфозом. Онтогенез закінчується утворенням дорослої тварини, готової до статевого розмноження.
У багатоклітинних є кілька типів дробіння яйця. Тип дробіння значною мірою залежить від кількості та локалізації жовтка в яйці. Якщо жовтка небагато й він розташований рівномірно, дробіння завершується повністю, з утворенням однакових бластомерів (повне, рівномірне дробіння). Якщо жовток сконцентрований на вегетативному полюсі, дробіння може відбуватися нерівномірно: на вегетативному полюсі утворюються великі, багаті на жовток макромери, на анімальному — дрібні мікромери. У разі збільшення кількості жовтка в яйці центральна маса цитоплазми з жовтком не ділиться, відбувається поверхневе неповне дробіння. При цьому ядро поділяється багаторазово, .дочірні ядра мігрують у периферійну частину цитоплазми, яка ділиться на клітини, а в центрі залишається жовток. У яєць з великою кількістю жовтка, що зосереджений на вегетативному полюсі, бластомери утворюються лише на анімальному полюсі, а вегетативна частина яйця не ділиться.
Типи дробіння також визначають за взаємним розташуванням бластомерів (рис. 63). Так, у .кишковопорожнинних і деяких плоских червів бластомери розташовані неупорядковано (анархічне дробіння). У голкошкірих відоме радіальне дробіння: перші дві борозни дробіння йдуть від полюса до полюса меридіональне, третя — по екватору; потім меридіональні та екваторіальні борозни чергуються, й бластомери розміщуються правильними горизонтальними та вертикальними рядами. Під час спірального дробіння мітотичні веретена розміщуються під кутом 45° до поздовжньої осі яйця — за годинниковою стрілкою або проти, причому в кожному наступному поділі напрямки поділу чергуються, В результаті бластомери розташовуються спіральне. При цьому, як правило, дробіння йде нерівномірно — кожен бластомер поділяється на макромер, локалізований ближче до вегетативного полюса, та мікромер, що міститься біля анімального полюса. Таке дробіння відоме, наприклад, у кільчастих червів.
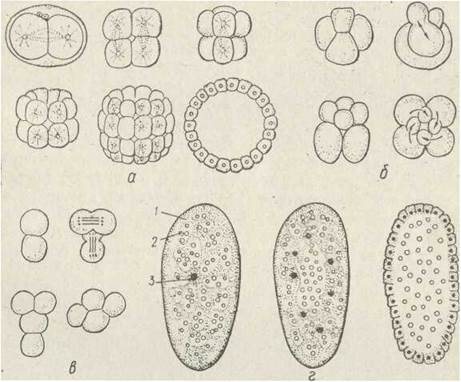
Рис. 63. Типи дробіння яйця — радіальне (а), спіральне (б), білатеральне (б), поверхневе (г):
1 — цитоплазма; 2 — гранули жовтка; 3 — ядро
У нематод має місце білатеральне дробіння, коли бластомери розподіляються симетрично з боків поздовжньої площини симетрії, яка зберігається й у дорослої тварини.
Дробіння закінчується стадією бластули, яка може мати різну будову, однак у більшості випадків має вигляд кулястого або сплюснутого утвору, який складається з одного шару клітин і всередині має більш-менш розвинену, заповнену рідиною порожнину бластоцель.
Далі утворюються зародкові листки — ектодерма, ентодерма та мезодерма, з яких формуються певні тканини дорослої тварини, Вчення про зародкові листки, що містяться в усіх багатоклітинних тварин (крім Prometazoa), було розроблено в XIX ст. рядом вчених — X. Пандером, К. Бером, О, О. Ковалевським, І. І. Мечниковим, Е. Геккелем.
Зародкові листки формуються таким чином. Спочатку утворюється двошаровий зародок — гаструла. Вона складається з ектодерми та ентодерми, яка встилає порожнину сліпо замкненої первинної кишки, що відкривається первинним ротом (бластопор) на вегетативному полюсі. Процес утворення гаструли -=- гаструляція — у різних тварин відбувається неоднаково. Розрізняють кілька типів гаструляції.
Імміграція характеризується тим, що частина клітин бластодерми мігрує в бластоцель, де утворює шар ендодерми. Імміграція буває мультиполярна, коли проходить но всій бластулі, та уніполярна, коли клітини мігрують лише на вегетативному полюсі. При цьому бластопор часто не виражений (рис 64).
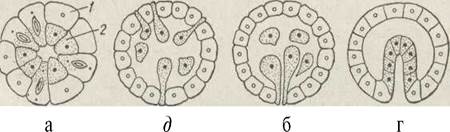
Рис. 64. Основні типи гаструляції - деламінація (а), мультиполярна імміграція (б), уніполярна імміграція (в), інвагінація (г):
1 — ектодерма; 2 — ентодерма
Деламінація полягає в тому, що клітини бластули поділяються навпіл паралельно поверхні бластули, утворюючи одразу екто- та ентодерму.
Інвагінація характеризується тим, що на вегетативному полюсі стінка вп'ячується й гаструла набуває вигляду двошарового мішка з бластопором і первинним кишечником, порожнина якого має назву гастроцель.
Епіболія полягає в тому, що великі клітини вегетативного полюса обростають дрібними клітинами анімального, при цьому гастроцель не утворюється, а бластопор має вигляд западини на вегетативному полюсі, куди не проникли дрібні клітини ектодерми.
Способи утворення третього зародкового листка — мезодерми досить різноманітні й розглядатимуться в розділах, де характеризуватимуться відповідні типи.
Розрізняють прямий постембріональний розвиток і з перетворенням. Під час прямого розвитку з яйця виходить особина, що відрізняється від дорослої лише розмірами та недорозвиненими статевими органами олігохети, п'явки, більшість турбелярій тощо). Розвиток з перетворенням характеризується різноманітними личинковими фазам, які відрізняються від дорослих за будовою та способом життя. У багатьох безхребетних із зовнішнім скелетом або товстою кутикулою (нематоди, членистоногі) ріст личинок супроводжується линьками.
Багатоклітинні тварини різняться між собою за рівнем організації, наявністю та кількістю зародкових листків, ступенем клітинної та органологічної диференціації, розвитком певних систем органів. Ми приймаємо поділ Metazoa на два розділи: Первинні багатоклітинні (Prometazoa) та Справжні багатоклітинні (Eumetazoa).