ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 1 - Г.Й. Щербак - 1995
ПІДЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
ПОХОДЖЕННЯ БАГАТОКЛІТИННИХ
Походження багатоклітинних досі остаточно не з'ясоване. Ще в минулому столітті вчені дискутували з приводу походження багатоклітинних, висуваючи різні, інколи навіть фантастичні, гіпотези. До нашого часу зберегли своє значення лише кілька з них, насамперед ті, де .визнається, що предками багатоклітинних були найпростіші. Ці гіпотези можна розділити на дві групи.
До першої групи належать гіпотези про походження багатоклітинних від колоній найпростіших. Так, у 70-х роках минулого століття відомий німецький біолог Е. Геккель розвинув систему поглядів на походження багатоклітинних від колоніальних джгутикових — гіпотезу гастреї. Згідно з цією гіпотезою предками багатоклітинних були кулясті колонії джгутикових, подібні до сучасних.
Геккель спирався на дані ембріології й надавав основним етапам ембріонального розвитку організму філогенетичного значення. Подібно до того, як у онтогенезі багатоклітинний організм утворюється з однієї заплідненої яйцеклітини, що в результаті дробіння перетворюється на багатоклітинні стадії — морулу, потім бластулу та гаструлу, так і в історичному розвитку — спочатку виникли одні неклітинні амебоподібні організми — цитеї, потім від таких організмів розвинулися колонії з кількох особин — мореї, які згодом перетворилися на кулясті одношарові колонії — бластеї, що мали на поверхні джгутики та плавали в товщі води. Нарешті, вп'ячування стінки бластеї всередину (інвагінація) привело до виникнення двошарового організму — гастреї. Зовнішній шар її клітин мав джгутики й виконував локомоторну функцію, а внутрішній вистилав первинний кишечник і виконував функцію травлення. Так, за гіпотезою Геккеля, одночасно виникли первинний рот (бластопор) і замкнена первинна кишка. Оскільки за часів створення цієї гіпотези єдиним способом гаструляції вважалася інвагінація, властива більш високоорганізованим тваринам (ланцетник, асцидії, щетинкощелепні), Геккель твердив, що й у філогенезі утворення багатоклітинних гастрей відбувалося саме таким чином. З двошарового плаваючого організму — гастреї, яка осіла на субстрат на аборальний полюс, почався розвиток кишковопорожнинних, що є, на думку Геккеля, найпримітивнішими багатоклітинними, від яких виникли всі інші багатоклітинні (рис. 65, а — в).
На свій час гіпотеза гастреї була достатньо обгрунтованою. Гекель висунув її ще до відкриття І. І Мечниковим внутрішньоклітинного травлення. Тоді вважалося, що їжа перетравлюється лише в порожнині кишечника, тому й первинну ентодерму уявляли у вигляді епітелію первинної кишки.
Гіпотеза гастреї відіграла велику роль у розвитку еволюційної зоології. У ній вперше було обгрунтовано єдність походження усіх багатоклітинних тварин. Гіпотезу підтримав ряд зоологів, з певними доповненнями її приймає й чимало сучасних вчених, зокрема в Західній Європі, вона викладена також у багатьох зарубіжних підручниках зоології.
Однак ще сучасники Геккеля зазначали недоліки гіпотези. Одним із найважливіших є відсутність обгрунтування, яке б пояснювало фізіологічні причини вп'ячування стінки бластеї всередину під час утворення гастреї. Гіпотеза гастреї не пояснює також існування в нижчих багатоклітинних личинки типу паренхімули (губки, нижчі кишковопорожнинні), яка не має епітелізованої ентодерми.
Однією з модифікацій гіпотези гастреї була гігіотеза плакули, запропонована англійським ученим О. Вючлі (1884), який вважав, що багатоклітинні походять із двошарової пласкої колонії найпростіших (плакули). Звернений до субстрату шар плакули виконував функцію живлення, поглинаючи харчові частки з дна. Вигинаючись одним боком догори, двошарова плакула перетворилася на гастреєподібний організм (рис. 65, г — е).
Досить популярною серед сучасних вчених є інша модифікація гіпотези гастреї, висунута шведським вченим Т. Єгерстеном у 1955—1972 рр. і відома під назвою гіпотези білатерогастреї. Згідно з цією гіпотезою віддаленим предком багатоклітинних тварин була куляста колонія рослинних джгутикових, схожа на Volvox, яка плавала в поверхневих шарах води й могла живитися автотрофне та гетеротрофне — за рахунок фагоцитозу дрібних органічних часток. Колонія, як і сучасний Volvox, мала передньо-задню полярність. На думку Єгерстена, така бластея перейшла до бентосного типу життя, осівши на дно боком, який став пласким.
Таким чином виникла донна білатеральносиметрична (така, через тіло якої можна провести одну площину симетрії, що поділяє її на дві дзеркально подібні половини) бластулоподібна тварина - білатеробластея (рис. 65, є — з). Оскільки освітленість на дні недостатня для фотосинтезу, білатеробластея живилася переважно гетеротрофне, фагоцитуючи поживні частки з дна клітинами вентрального епітелію. Під час переходу до, живлення крупнішою
здобиччю ці тварини втягували вентральний шар, утворюючи тимчасову порожнину, до якої потрапляла здобич і де відбувалося її перетравлення. Поступово така тимчасова порожнина стала постійною кишковою порожниною., яка відкривалася назовні щілиноподібним, витягнутим у передньо-задньому напрямку ротом. Так виникла білатерогастрея - гаструлоподібний білатеральносиметричний організмів.
Від білатерогастреї походять рубки, які, на думку Єгерстена, мають кишкову порожнину. Пізніше, в процесі еволюції білатерогастреї з'явилися три пари бічних вп'ячувань у стінках кишечника. Від такої ускладненої білатерогастреї походять усі інші типи тварин: кишковопорожнинні (первинні коралові поліпи) з трьома парами септ у гастральній порожнині, а також целомічні тварини з трьома парами целомів. Паренхімні та первиннопорожнинні тварини, за цією гіпотезою, вторинно втратили целом.
Гіпотези плакули та білатерогастреї мають ті ж недоліки, що й класична гіпотеза гастреї — вони припускають, що первинною вихідною гаструлою е саме інвагінаційна, яка у нижчих багатоклітинних трапляється дуже рідко. До того ж гіпотеза білатерогастреї базується на тому, що первинним типом симетрії в багатоклітинних була білатеральна, а не суперечить ембріологічним відомостям, за якими в. онтогенезі нижчих багатоклітинних немає навіть -її слідів. Палеонтологічні дослідження свідчать, що ці типи симетрії могли виникнути в різних Metazoa одночасно та незалежно.
Нині найбільш обгрунтованою й альтернативною гіпотезі гастреї можна вважати гіпотезу вітчизняного вченого І. І. Мечникова, розроблену в 1877—1886 рр. Вивчаючи ембріональний розвиток нижчих багатоклітинних — губок і кишковопорожнинних, Мечников встановив, що в процесі утворення двошарової стадії в них відбувається не впинання, а здебільшого імміграція — вповзання окремих клітин стінки бластули до її порожнини. Цей примітивний процес утворення гаструли Мечников вважав первинним, а інвагінацію — наслідком скорочення та спрощення розвитку, що мали місце в процесі еволюції.
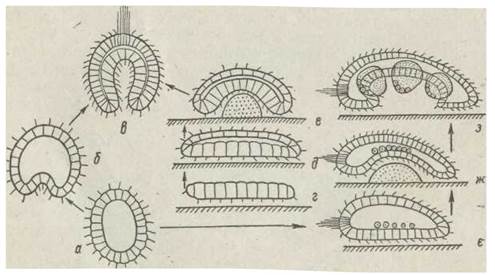
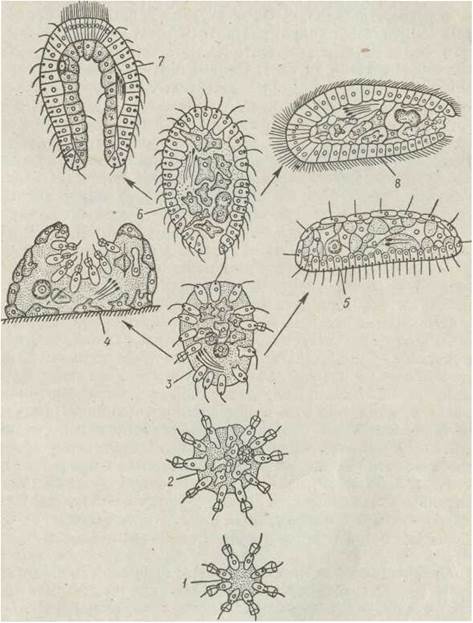
Рис. 65. Походження багатоклітинних — теорія гастреї (а—в), плакули (г—е), білатерогастреї (є—з), фагоцители (й): 2 — колонії хоанофлагелят; 3 — рання фагоцитела (гіпотетичний предок Prometazoa); 4— примітивна губка; 5 — Placozoa; 6 — пізня фагоцитела з ротовим отвором (гіпотетичний предок Enmetazoa); 7 — гастрея (гіпотетичний предок кишковопорожнинних); 8— первинна турбелярія
Предками багатоклітинних, за гіпотезою Мечникова, бути кулясті колонії гетеротрофних джгутикових, які плавали у воді, живилися, фагоцитуючи дрібні частки, окремими клітинами колонії/ Прототипом такої колонії можуть були пелагічні кулясті колонії комірцевих джгутикових (Sphaeroeca volvox). Окремі клітини, захопивши поживну часточку, втрачали джгутик, перетворюючись на амебоїдних, і занурювалися вглиб колонії, заповненої безструктурною драглистою речовиною. Потім вони могли повертатися на поверхню. Таке явище спостерігається в сучасних губок, джгутикові клітини хоаноцити яких можуть, заповнившись їжею, перетворюватися на амебоїдні й мігрувати до паренхіми, де відбувається травлення, а потім повертатися на місце. З часом клітини диференціювалися на ті, що забезпечували переважно рух колонії, й ті, які живилися та годували інших. Колонія вже не мала вигляду порожньої кулі — всередині містилося скупчення фагоцитів. З сучасних тварин до організмів такого типу найближчими є комірцеві джгутикові (ряд Choanoflagellida) Proterospongia haeckeli, що утворюють колонію, в зовнішньому шарі якої містяться комірцеві джгутикові, а у внутрішньому — амебоїдні клітини. Поступово тимчасове диференціювання клітин набуло постійного характеру й колонія одноклітинних перетворилася на багатоклітинний організм, який мав два шари клітин: зовнішній (джгутиковий) — кінобласт і внутрішній (амебоїдний) — фагоцитобласт. Живлення такого організму відбувалося за рахунок захоплення джгутиковими клітинами кінобласта органічних часток із товщі води й передачі їх амебоїдним клітинам фагоцитобласта. Цей гіпотетичний багатоклітинний організм Мечников назвав фагоцителою, бажаючи підкреслити роль фагоцитозу в його виникненні (див. рис. 65, 66).
Організація фагоцители близька до будови личинок губок і гідроїдних — паренхімул. Саме під час формування цих личинок відбувається міграція клітин із поверхні бластули до її порожнини. Від таких фагоцителоподібних предків, на думку Мечникова, походять губки та кишковопорожнинні.
Гіпотеза Мечникова спиралася на великий ембріологічний матеріал і дослідження з фізіології. Над подальшим ствердженням і обгрунтуванням гіпотези фагоцители працювали видатні російські зоологи О. О. Захваткін (1949), В. М. Беклемішев (1944, 1964), А. В. Іванов (1968), англійська дослідниця Л. Гайман (1940, 1951). Ці вчені збагатили її новими доказами, одержаними внаслідок цитологічних, ембріологічних і протозоологічних досліджень, перетворивши на науково обгрунтовану теорію.
О. О. Захваткін, який вивчав сучасних колоніальних найпростіших, показав принципову можливість перетворення колонії на цілісний багатоклітинний організм. Проте він вважав, що фагоцители — це не дорослі організми, а вільноплаваючі личинки, які не живилися й не розмножувалися, а лише розселялися. Дорослі стадії, на думку Захваткіна, були сидячими, прикріпленими до дна колоніальними організмами, схожими на сучасних губок і кишковопорожнинних. Первинні багатоклітинні мали складний життєвий цикл, подібний до метагенезу (Hydrozoa) з чергуванням нестатевих поліпоїдних поколінь і статевого покоління медуз.
Багато нового вніс у теорію фагоцители А. В. Іванов. Він не лише навів нові цитологічні та ембріологічні докази на її користь, а й показав, якими за будовою та життєвим циклом мали бути предки Metazoa. Розвиваючи теорію фагоцители, А. В. Іванов описав зміни, які могли відбутися в фагоцителоподібних тварин у процесі їх подальшої еволюції у разі збереження плаваючого способу життя й переходу до сидячого або повзаючого способу життя. Одні нащадки фагоцители перейшли від плаваючого способу життя до сидячого (саме від них походять губки, які ще не мають рота, кишечника, нервової системи та мускулатури), інші набули здатності повзати по дну, їх кінобласт диференціювався на вентральний і дорзальний епітелії. Від них походять Placozoa (Тгіchoplax) які, за сучасними поглядами, є реліктовими тваринами, що залишилися на рівні організації фагоцители. Серед сучасних тварин на такому ж рівні організації перебувають Dicyemida та Orthonectida.
Частина фагоцителоподібних тварин зберегла плаваючий спосіб життя, й у процесі еволюції в них з'явилися рот, нервова система, м'язи. Від таких розвинених фагоцител виникли,, з одного боку, первинні кишковопорожнинні, що перетворилися на сидячі поліпи, а з іншого — повзаючі по дну предки сучасних безкишкових турбелярій, які набули -білатеральної симетрії, але ще не мали кишечника — від них походять плоскі черви (див. рис. 65).
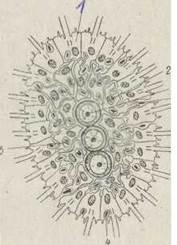
Рис. 66. Схема організації фагоцители:
1 — кінобласт; 2 — фагоцитобласт; 3 — статеві клітини; 4 — фагоцитоз; 5 — травна вакуоля
До другої групи гіпотез походження багатоклітинних належать різні модифікації гіпотези целюляризації. її прихильники виходять з того, що одноклітинне найпростіше відповідає не окремій клітині багатоклітинного, а цілому багатоклітинному організму. Целюляризація — це розпад на окремі клітини. Вперше ідею целюляризації ще наприкінці минулого століття висунули Г. Нерінг (1877), І. Деляж (1896). У XX ст. цю ідею розвивали. Й. Хаджі (1944, 1963) та інші вчені. На думку цих вчених, багатоклітинні походять від багатоядерних інфузорій. Перехід від одноклітинного стану до багатоклітинного відбувався в тілі інфузорій одночасно шляхом утворення клітинних меж навколо окремих ядер і прилеглих до них ділянок цитоплазми — енергід.
Складно побудована клітина інфузорій нібито перетворилася на досить високоорганізованого черва (турбелярію) або навіть коловертку, органи якого утворюються з її органел. Рот і глотка сформувалися з цитостома та цитофаринкса, кишка — з ендоплазми, анус — із порошиці, видільні органи — із скоротливих вакуолей, статева система—з мікронуклеусів. Ця концепція має своїх прихильників серед вчених різних країн. Проте важко уявити, як спеціалізовані, складно побудовані інфузорії, ядерний апарат (ядерний дуалізм), диференціація покривів (складний кортекс), особливості статевого процесу (кон'югація) та життєвого циклу яких не мають нічого спільного з будовою та функціями інших тварин, могли дати початок багатоклітинним тваринам. Найбільше заперечень викликає припущення прихильників гіпотези целюляризації про можливість перетворення спеціалізованих органел і ділянок клітини на багатоклітинні тканини та органи з тими ж функціями. Ембріологічні відомості також суперечать гіпотезі целюляризації. Більше того, за цією гіпотезою, тварини, що стоять нижче коловерток і турбелярій, виникли внаслідок регресивної еволюції.
Усі наведені гіпотези виходять з того, що багатоклітинні мають монофілетичне походження, тобто усі вони тим або іншим шляхом виникли від однієї предкової форми (гіпотетичної гастреї, планули, білатерогастреї, фагоцители або інфузорії). Проте останнім часом з'являється
.дедалі більше доказів, що губки мають незалежне від інших Metazoa походження. В результаті електронно-мікроскопічного порівняльного дослідження будови хоаноцитів губок і хоанофлагелят виявлено майже повну їх ідентичність, з одного боку, та істотні відмінності від комірцево- джгутикових клітин інших багатоклітинних — з іншого. Такі клітини є, наприклад, у епітелії «апівхордових, деяких голкошкірих тощо. Цей факт, а також велика своєрідність губок (відсутність у них рота, кишечника, нервів, м'язів, зародкових листків) свідчать про те, що губки ви- виникли від стародавніх хоанофлагелят або спільних з ними предків, а всі інші багатоклітинні —від якихось гетеротрофних колоніальних джгутикових, що до нашого часу не дожили. Якщо дотримуватися цієї точки зору, слід погодитися з тим, що предки губок і предки інших Metazoa — це різні тварини, але будова їх могла бути подібною. Адже паренхімули губок і кишковопорожнинних однакові, хоча, можливо, й мають різне походження.